

Specific encapsidation of nodavirus RNAs is mediated through the C terminus of capsid precursor protein alpha
- PMID: 9765417
- PMCID: PMC110289
- DOI: 10.1128/JVI.72.11.8738-8746.1998
Specific encapsidation of nodavirus RNAs is mediated through the C terminus of capsid precursor protein alpha
Abstract
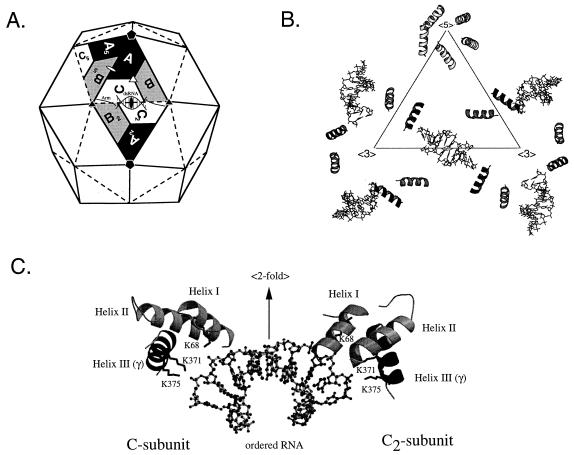
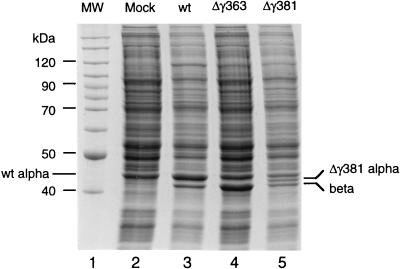
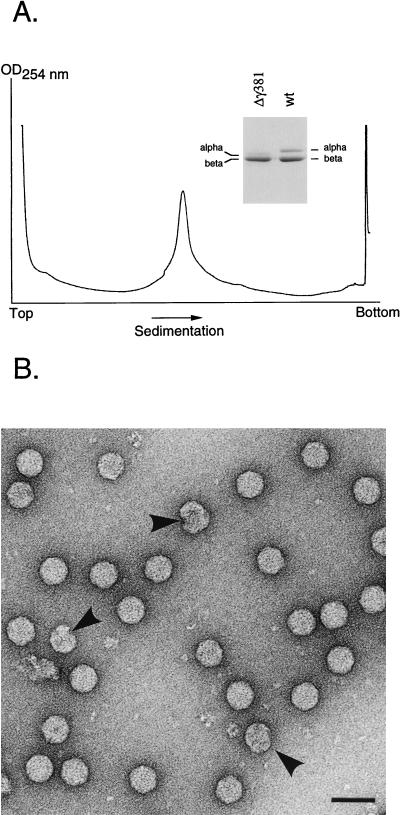
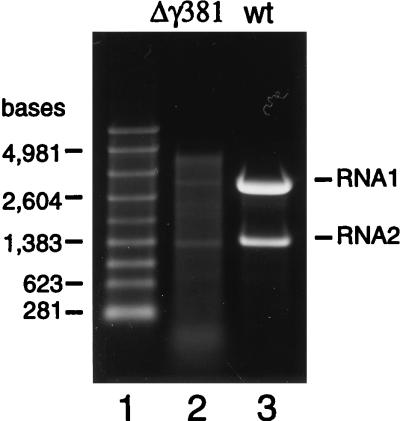
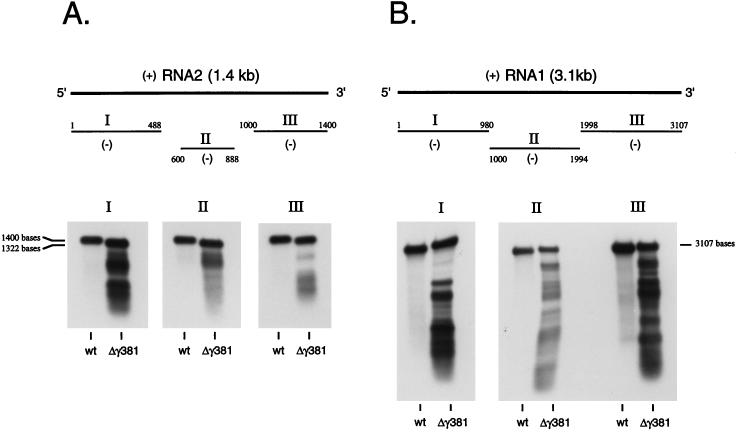
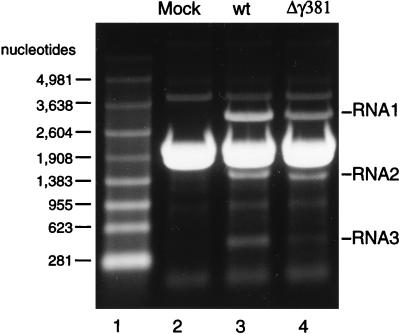
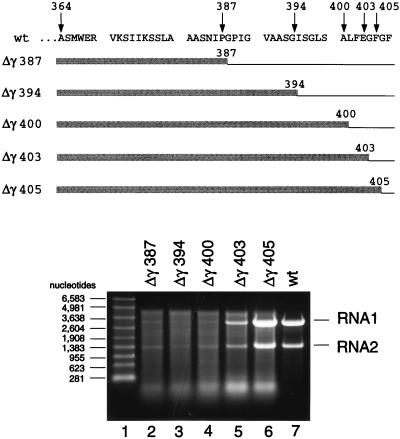
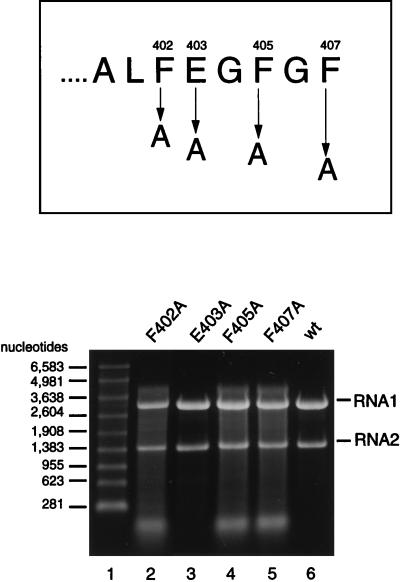
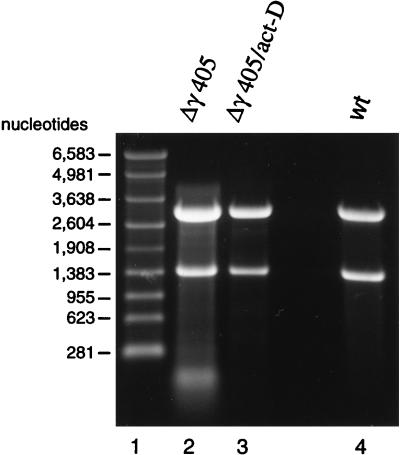
Flock house virus (FHV) is a small icosahedral insect virus with a bipartite, messenger-sense RNA genome. Its T=3 icosahedral capsid is initially assembled from 180 subunits of a single type of coat protein, capsid precursor protein alpha (407 amino acids). Following assembly, the precursor particles undergo a maturation step in which the alpha subunits autocatalytically cleave between Asn363 and Ala364. This cleavage generates mature coat proteins beta (363 residues) and gamma (44 residues) and is required for acquisition of virion infectivity. The X-ray structure of mature FHV shows that gamma peptides located at the fivefold axes of the virion form a pentameric helical bundle, and it has been suggested that this bundle plays a role in release of viral RNA during FHV uncoating. To provide experimental support for this hypothesis, we generated mutant coat proteins that carried deletions in the gamma region of precursor protein alpha. Surprisingly, we found that these mutations interfered with specific recognition and packaging of viral RNA during assembly. The resulting particles contained large amounts of cellular RNAs and varying amounts of the viral RNAs. Single-site amino acid substitution mutants showed that three phenylalanines located at positions 402, 405, and 407 of coat precursor protein alpha were critically important for specific recognition of the FHV genome. Thus, in addition to its hypothesized role in uncoating and RNA delivery, the C-terminal region of coat protein alpha plays a significant role in recognition of FHV RNA during assembly. A possible link between these two functions is discussed.
Figures

References
-
- Convery M A, Rowsell S, Stonehouse N J, Ellington A D, Murray H I J B, Peabody D S, Phillips S E V, Stockley P G. Crystal structure of an aptamer-protein complex at 2.8 A resolution. Nat Struct Biol. 1998;5:133–139. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
