

Induction of programmed cell death by parvovirus H-1 in U937 cells: connection with the tumor necrosis factor alpha signalling pathway
- PMID: 9765434
- PMCID: PMC110306
- DOI: 10.1128/JVI.72.11.8893-8903.1998
Induction of programmed cell death by parvovirus H-1 in U937 cells: connection with the tumor necrosis factor alpha signalling pathway
Abstract
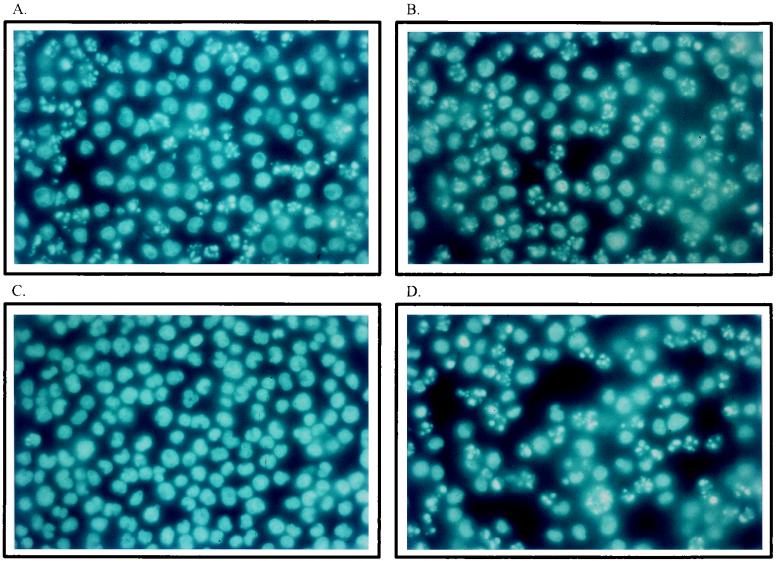
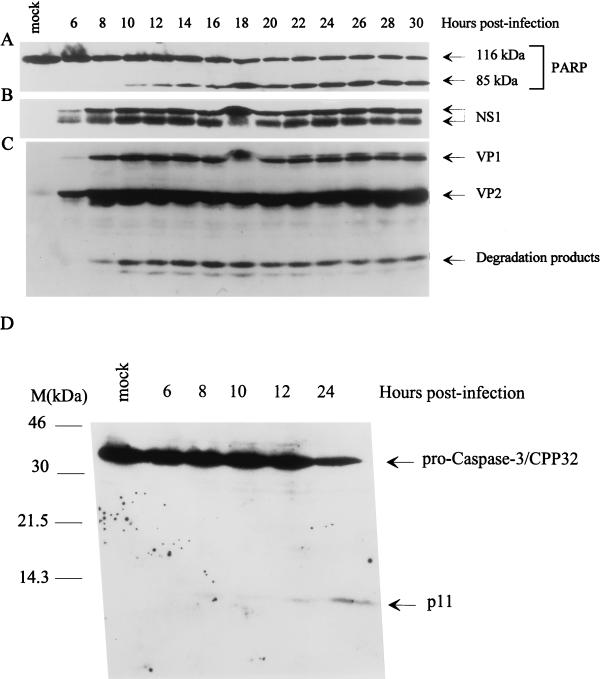
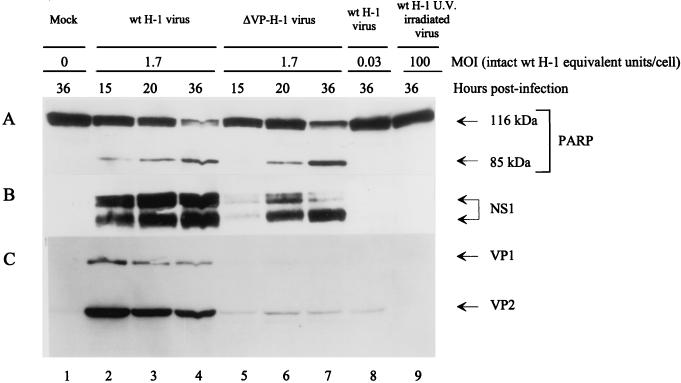
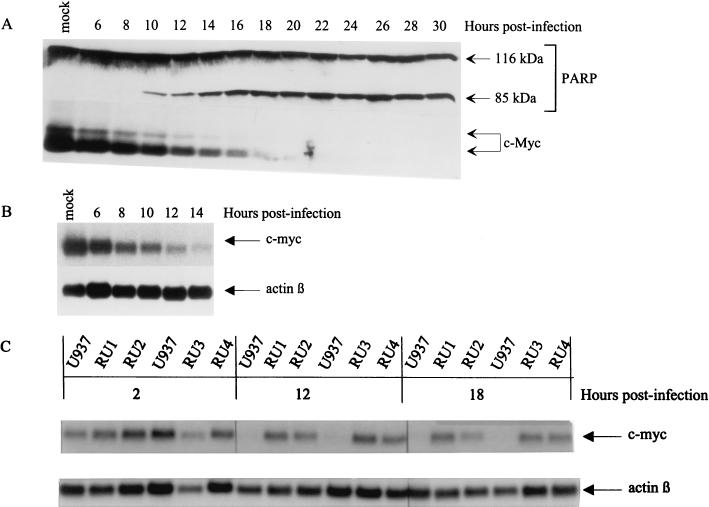
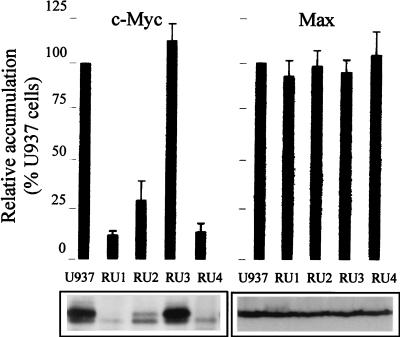
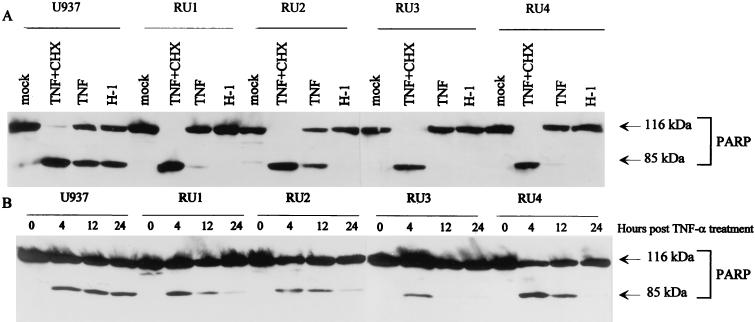
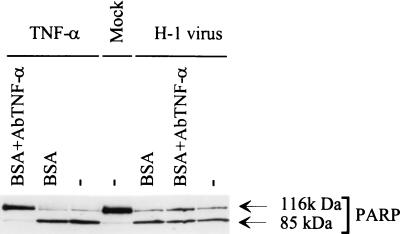
The human promonocytic cell line U937 undergoes apoptosis upon treatment with tumor necrosis factor alpha (TNF-alpha). This cell line has previously been shown to be very sensitive to the lytic effect of the autonomous parvovirus H-1. Parvovirus infection leads to the activation of the CPP32 ICE-like cysteine protease which cleaves the enzyme poly(ADP-ribose)polymerase and induces morphologic changes that are characteristic of apoptosis in a way that is similar to TNF-alpha treatment. This effect is also observed when the U937 cells are infected with a recombinant H-1 virus which expresses the nonstructural (NS) proteins but in which the capsid genes are replaced by a reporter gene, indicating that the induction of apoptosis can be assigned to the cytotoxic nonstructural proteins in this cell system. The c-Myc protein, which is overexpressed in U937 cells, is rapidly downregulated during infection, in keeping with a possible role of this product in mediating the apoptotic cell death induced by H-1 virus infection. Interestingly, four clones (designated RU) derived from the U937 cell line and selected for their resistance to H-1 virus (J. A. Lopez-Guerrero et al., Blood 89:1642-1653, 1997) failed to decrease c-Myc expression upon treatment with differentiation agents and also resisted the induction of cell death after TNF-alpha treatment. Our data suggest that the RU clones have developed defense strategies against apoptosis, either by their failure to downregulate c-Myc and/or by activating antiapoptotic factors.
Figures
References
-
- Amati B, Land H. Myc-Max-Mad: a transcription factor network controlling cell cycle progression, differentiation and death. Curr Opin Genet Dev. 1994;4:102–108. - PubMed
-
- Askew D S, Ashmun R A, Simmons B C, Cleveland J L. Constitutive c-myc expression in an IL-3-dependent myeloid cell line suppresses cell cycle arrest and accelerates apoptosis. Oncogene. 1991;6:1915–1922. - PubMed
-
- Beg A A, Baltimore D. An essential role for NF-κB in preventing TNF-α-induced cell death. Science. 1996;274:782–784. - PubMed
-
- Beidler D R, Tewari M, Friesen P D, Poirier G, Dixit V M. The baculovirus p35 protein inhibits Fas- and tumor necrosis factor-induced apoptosis. J Biol Chem. 1995;270:16526–16528. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
