

The 5' and 3' TAR elements of human immunodeficiency virus exert effects at several points in the virus life cycle
- PMID: 9765469
- PMCID: PMC110341
- DOI: 10.1128/JVI.72.11.9217-9223.1998
The 5' and 3' TAR elements of human immunodeficiency virus exert effects at several points in the virus life cycle
Abstract
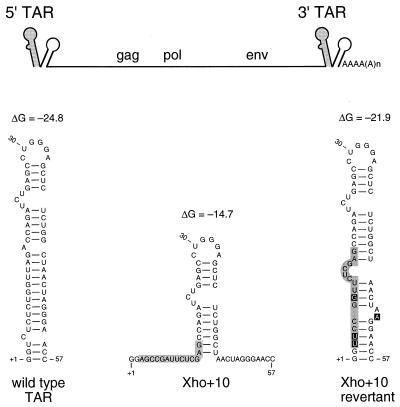
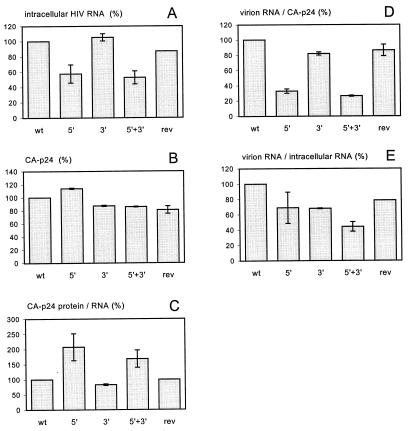
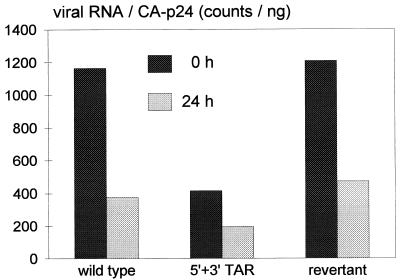
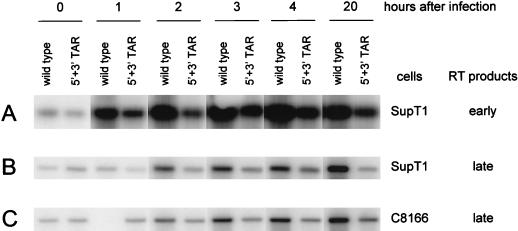
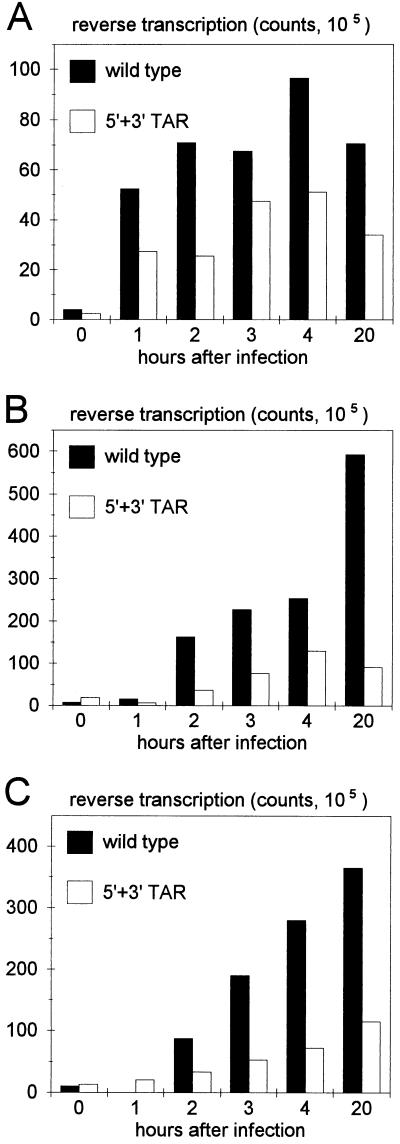
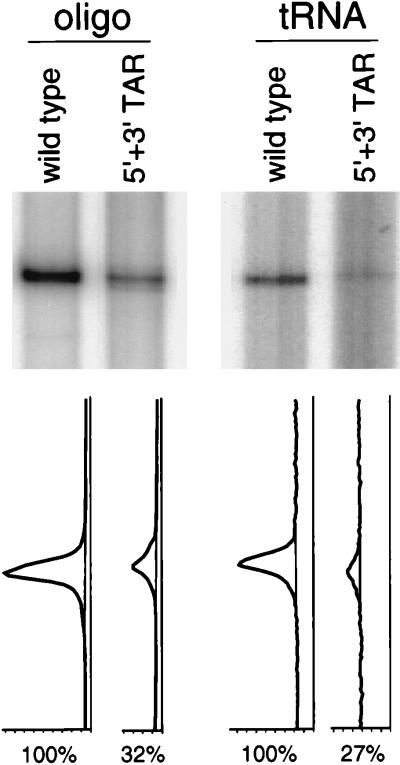
The human immunodeficiency virus type 1 RNA genome contains a terminal repeat (R) sequence that encodes the TAR hairpin motif, which has been implicated in Tat-mediated activation of transcription. More recently, a variety of other functions have been proposed for this structured RNA element. To determine the replicative roles of the 5' and 3' TAR hairpins, we analyzed multiple steps in the life cycle of wild-type and mutant viruses. A structure-destabilizing mutation was introduced in either the 5', the 3', or both TAR motifs of the proviral genome. As expected, opening of the 5' TAR hairpin caused a transcription defect. Because the level of protein expression was not similarly reduced, the translation of this mRNA was improved. No effect of the 3' hairpin on transcription and translation was measured. Mutations of the 5' and 3' hairpin structures reduced the efficiency of RNA packaging to similar extents, and RNA packaging was further reduced in the 5' and 3' TAR double mutant. Upon infection of cells with these virions, a reduced amount of reverse transcription products was synthesized by the TAR mutant. However, no net reverse transcription defect was observed after correction for the reduced level of virion RNA. This result was confirmed in in vitro reverse transcription assays. These data indicate that the 5' and 3' TAR motifs play important roles in several steps of the replication cycle, but these structures have no significant effect on the mechanism of reverse transcription.
Figures
References
-
- Arts E J, Li X, Gu Z, Kleiman L, Parniak M A, Wainberg M A. Comparison of deoxyoligonucleotide and tRNA(Lys-3) as primers in an endogenous human immunodeficiency virus-1 in vitro reverse transcription/template-switching reaction. J Biol Chem. 1994;269:14672–14680. - PubMed
-
- Auersperg N. Long-term cultivation of hypodiploid human tumor cells. J Natl Cancer Inst. 1964;32:135–163. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
