

Purification and characterization of NADP+-linked isocitrate dehydrogenase from scots pine . Evidence for different physiological roles of the enzyme in primary development
- PMID: 9765548
- PMCID: PMC34838
- DOI: 10.1104/pp.118.2.617
Purification and characterization of NADP+-linked isocitrate dehydrogenase from scots pine . Evidence for different physiological roles of the enzyme in primary development
Abstract
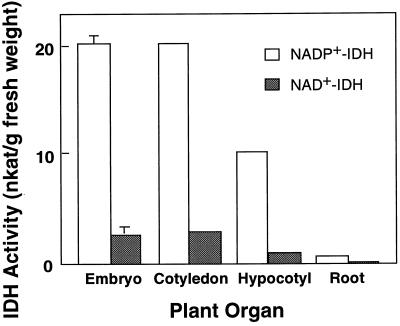
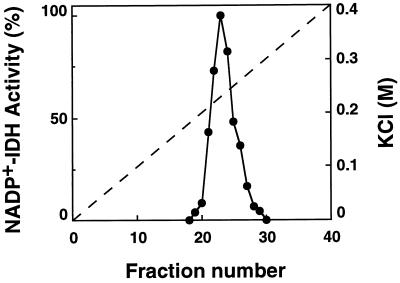
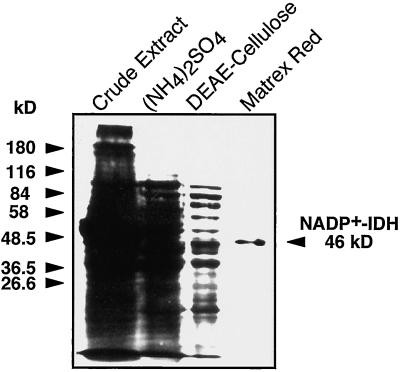
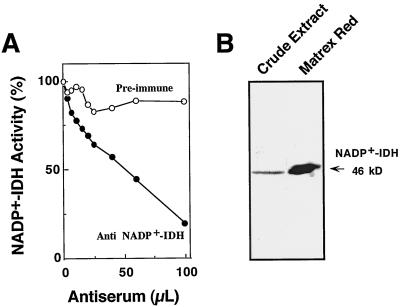
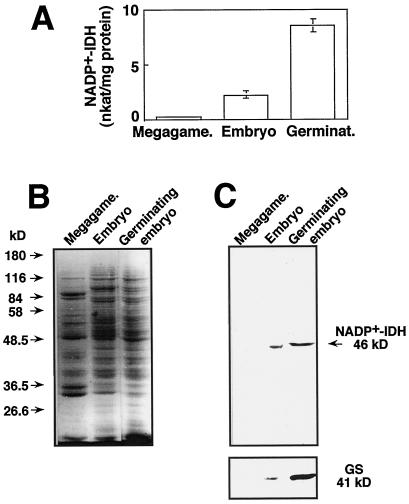
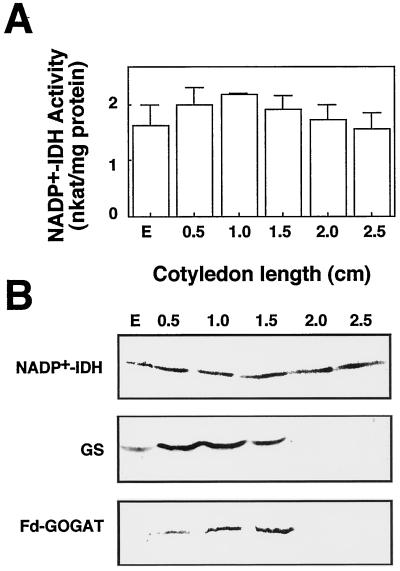
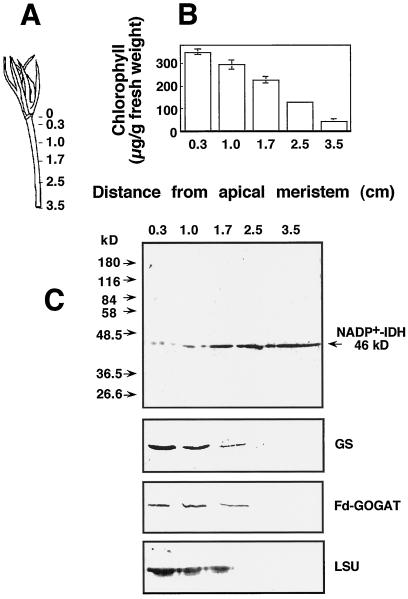
NADP+-isocitrate dehydrogenase (NADP+-IDH; EC 1.1.1.42) is involved in the supply of 2-oxoglutarate for ammonia assimilation and glutamate synthesis in higher plants through the glutamine synthetase/glutamate synthase (GS/GOGAT) cycle. Only one NADP+-IDH form of cytosolic localization was detected in green cotyledons of pine (Pinus spp.) seedlings. The pine enzyme was purified and exhibited molecular and kinetic properties similar to those described for NADP+-IDH from angiosperm, with a higher catalytic efficiency (10(5) M-1 s-1) than the deduced efficiencies for GS and GOGAT in higher plants. A polyclonal antiserum was raised against pine NADP+-IDH and used to assess protein expression in the seedlings. Steady-state levels of NADP+-IDH were coordinated with GS during seed germination and were associated with GS/GOGAT enzymes during chloroplast biogenesis, suggesting that NADP+-IDH is involved in the provision of carbon skeletons for the synthesis of nitrogen-containing molecules. However, a noncoordinated pattern of NADP+-IDH and GS/GOGAT was observed in advanced stages of cotyledon development and in the hypocotyl. A detailed analysis in hypocotyl sections revealed that NADP+-IDH abundance was inversely correlated with the presence of GS, GOGAT, and ribulose-1,5-bisphosphate carboxylase/oxygenase but was associated with the differentiation of the organ. These results cannot be explained by the accepted role of the enzyme in nitrogen assimilation and strongly suggest that NADP+-IDH may have other, as-yet-unknown, biological functions.
Figures

References
-
- Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem. 1976;72:248–254. - PubMed
-
- Bray CM. Nitrogen Metabolism in Plants. London: Longman; 1983.
-
- Canino S, Nieri B, Pistelli L, Alpi A, De Bellis L. NADP+-isocitrate dehydrogenase in germinating cucumber cotyledons: purification and characterization of a cytosolic isoenzyme. Physiol Plant. 1996;98:13–19.
LinkOut - more resources
Full Text Sources
Miscellaneous
