

Regulation of the Bacillus subtilis GlcT antiterminator protein by components of the phosphotransferase system
- PMID: 9765562
- PMCID: PMC107579
- DOI: 10.1128/JB.180.20.5319-5326.1998
Regulation of the Bacillus subtilis GlcT antiterminator protein by components of the phosphotransferase system
Abstract
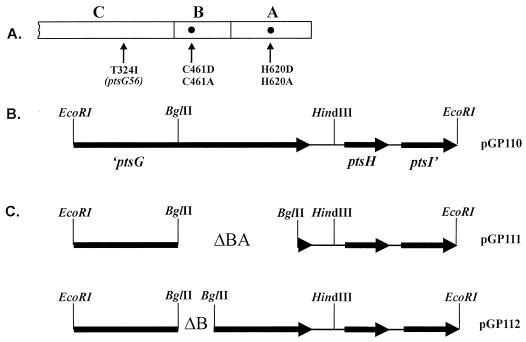
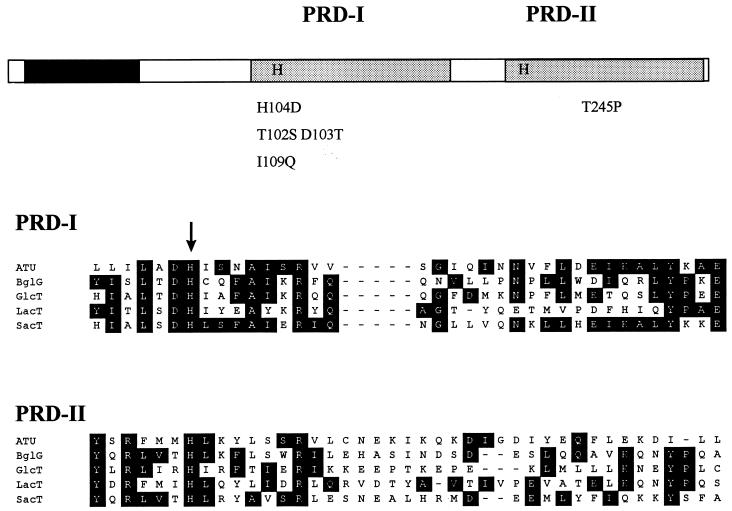
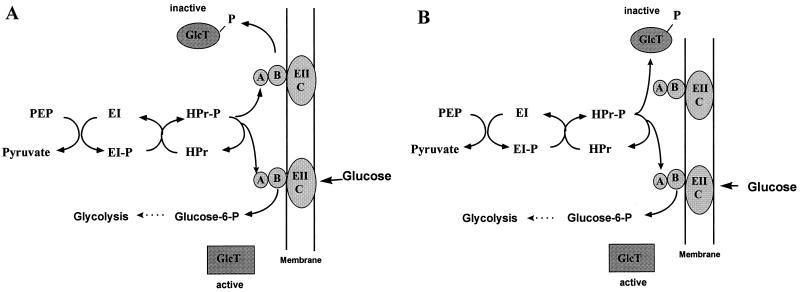
Bacillus subtilis utilizes glucose as the preferred source of carbon and energy. The sugar is transported into the cell by a specific permease of the phosphoenolpyruvate:sugar phosphotransferase system (PTS) encoded by the ptsGHI operon. Expression of this operon is induced by glucose and requires the action of a positive transcription factor, the GlcT antiterminator protein. Glucose availability is sensed by glucose-specific enzyme II (EIIGlc), the product of ptsG. In the absence of inducer, the glucose permease negatively controls the activity of the antiterminator. The GlcT antiterminator has a modular structure. The isolated N-terminal part contains the RNA-binding protein and acts as a constitutively acting antiterminator. GlcT contains two PTS regulation domains (PRDs) at the C terminus. One (PRD-I) is the target of negative control exerted by EIIGlc. A conserved His residue (His-104 in GlcT) is involved in inactivation of GlcT in the absence of glucose. It was previously proposed that PRD-containing transcriptional antiterminators are phosphorylated and concomitantly inactivated in the absence of the substrate by their corresponding PTS permeases. The results obtained with B. subtilis glucose permease with site-specific mutations suggest, however, that the permease might modulate the phosphorylation reaction without being the phosphate donor.
Figures
References
-
- Amster-Choder O, Houman F, Wright A. Protein phosphorylation regulates transcription of the β-glucoside utilization operon in E. coli. Cell. 1989;58:847–855. - PubMed
-
- Arnaud M, Débarbouillé M, Rapoport G, Saier M H, Jr, Reizer J. In vitro reconstitution of transcriptional antitermination by the SacT and SacY proteins of Bacillus subtilis. J Biol Chem. 1996;271:18966–18972. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
