

Regulation and function of T-cell-mediated immunity during Toxoplasma gondii infection
- PMID: 9767056
- PMCID: PMC88897
- DOI: 10.1128/CMR.11.4.569
Regulation and function of T-cell-mediated immunity during Toxoplasma gondii infection
Abstract
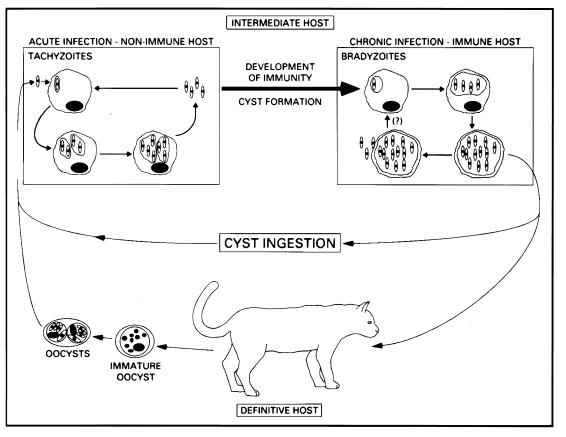
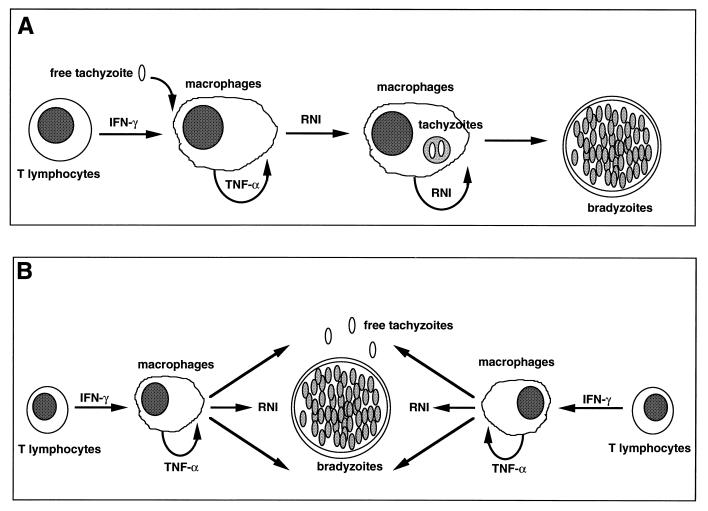
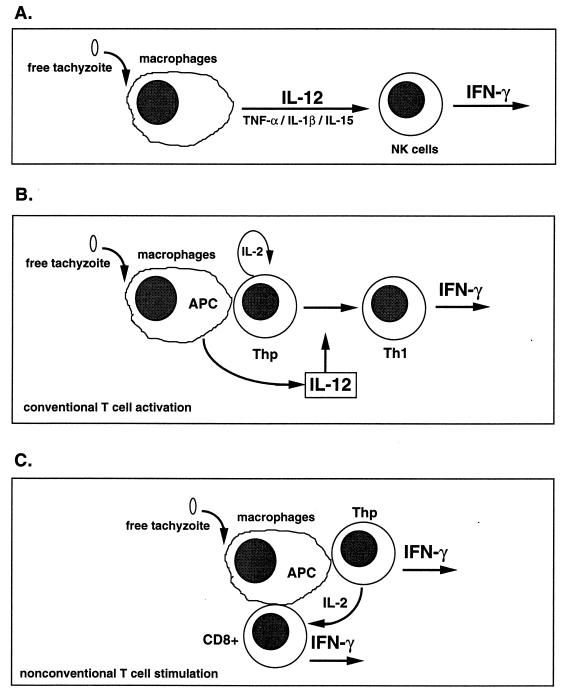
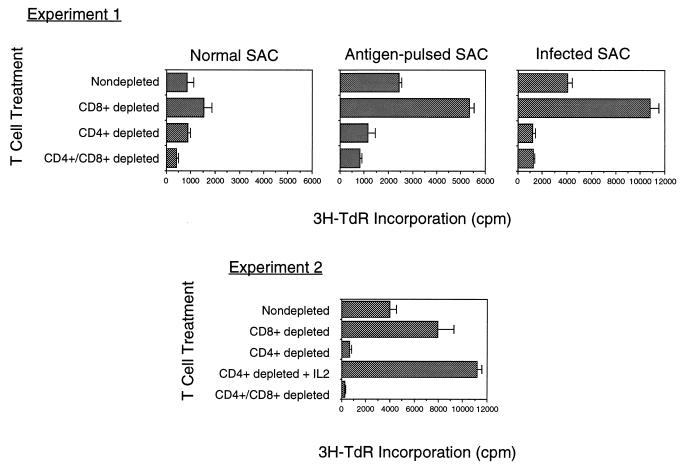
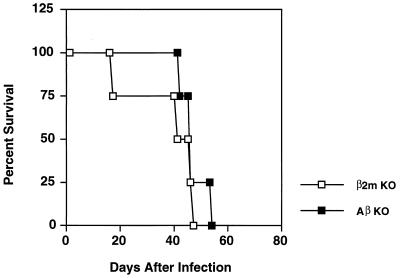
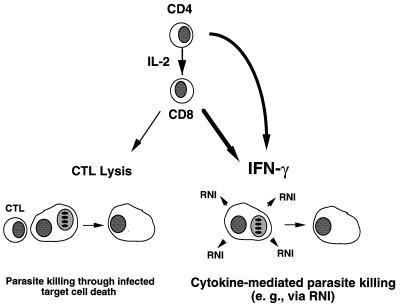
The intracellular protozoan Toxoplasma gondii is a widespread opportunistic parasite of humans and animals. Normally, T. gondii establishes itself within brain and skeletal muscle tissues, persisting for the life of the host. Initiating and sustaining strong T-cell-mediated immunity is crucial in preventing the emergence of T. gondii as a serious pathogen. The parasite induces high levels of gamma interferon (IFN-gamma) during initial infection as a result of early T-cell as well as natural killer (NK) cell activation. Induction of interleukin-12 by macrophages is a major mechanism driving early IFN-gamma synthesis. The latter cytokine, in addition to promoting the differentiation of Th1 effectors, is important in macrophage activation and acquisition of microbicidal functions, such as nitric oxide release. During chronic infection, parasite-specific T lymphocytes release high levels of IFN-gamma, which is required to prevent cyst reactivation. T-cell-mediated cytolytic activity against infected cells, while easily demonstrable, plays a secondary role to inflammatory cytokine production. While part of the clinical manifestations of toxoplasmosis results from direct tissue destruction by the parasite, inflammatory cytokine-mediated immunopathologic changes may also contribute to disease progression.
Figures
References
-
- Amichay D, Gazzinelli R T, Karupiah G, Moench T, Sher A, Farber J. The genes for the chemokines MuMig and Crg-2 are induced in protozoan and viral infections in response to IFN-γ with patterns of tissue expression that suggest nonredundant roles in vivo. J Immunol. 1996;157:4511–4520. - PubMed
-
- Aosai F, Yang T-H, Ueda M, Yano A. Isolation of naturally processed peptides from a Toxoplasma gondii-infected human B lymphoma cell line that are recognized by cytotoxic T lymphocytes. J Parasitol. 1994;80:260–266. - PubMed
-
- Apt W B. Toxoplasmosis in developing countries. Parasitol Today. 1985;1:44–46. - PubMed
-
- Araujo F G. Depletion of CD4+ T cells but not inhibition of the protective activity of IFN-γ prevents cure of toxoplasmosis mediated by drug therapy in mice. J Immunol. 1992;149:3003–3007. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
