

Inhibition of protease-resistant prion protein formation by porphyrins and phthalocyanines
- PMID: 9770449
- PMCID: PMC22794
- DOI: 10.1073/pnas.95.21.12117
Inhibition of protease-resistant prion protein formation by porphyrins and phthalocyanines
Abstract
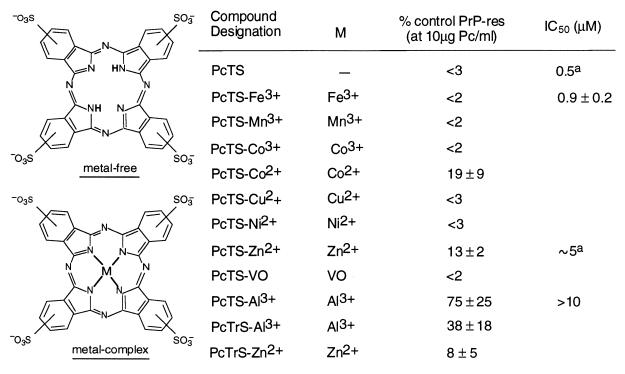
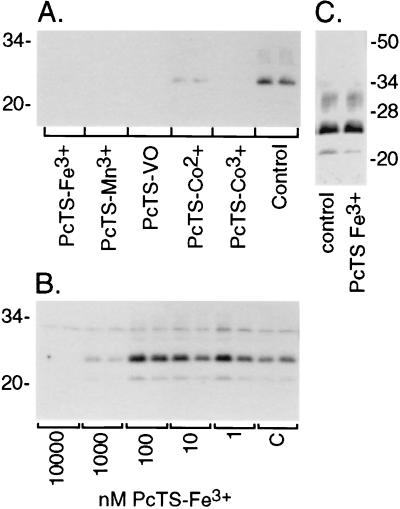
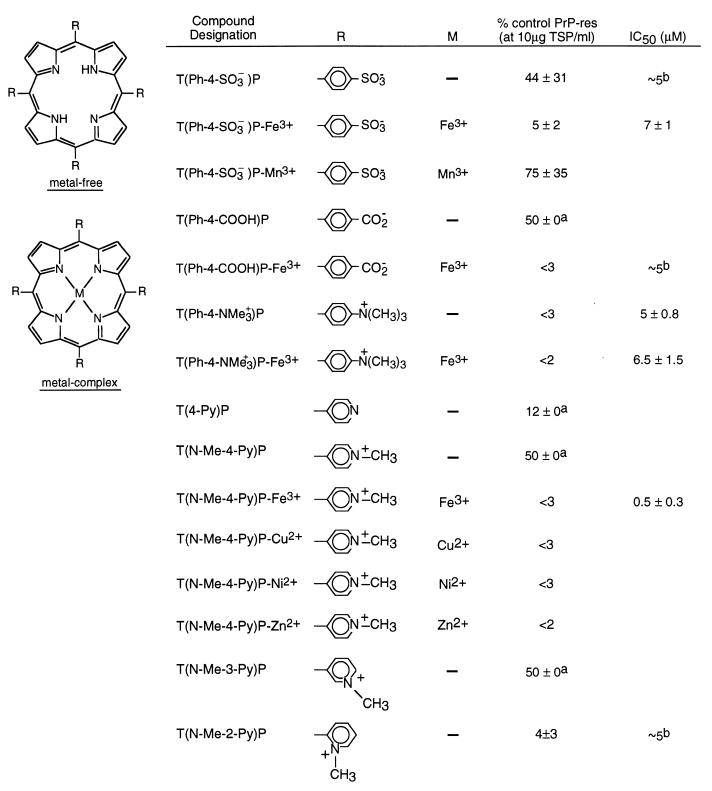
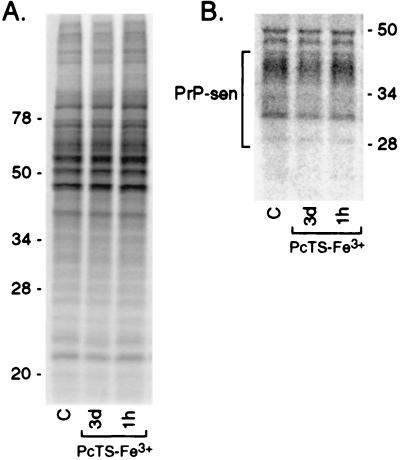
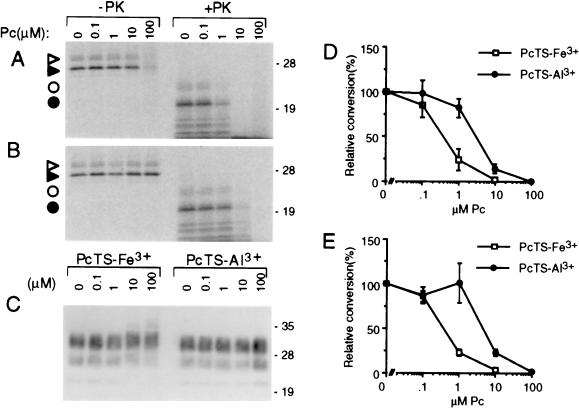
A central aspect of pathogenesis in the transmissible spongiform encephalopathies or prion diseases is the conversion of normal protease-sensitive prion protein (PrP-sen) to the abnormal protease-resistant form, PrP-res. Here we identify porphyrins and phthalocyanines as inhibitors of PrP-res accumulation. The most potent of these tetrapyrroles had IC50 values of 0.5-1 microM in scrapie-infected mouse neuroblastoma (ScNB) cell cultures. Inhibition was observed without effects on protein biosynthesis in general or PrP-sen biosynthesis in particular. Tetrapyrroles also inhibited PrP-res formation in a cell-free reaction composed predominantly of hamster PrP-res and PrP-sen. Inhibitors were found among phthalocyanines, deuteroporphyrins IX, and meso-substituted porphines; examples included compounds containing anionic, neutral protic, and cationic peripheral substituents and various metals. We conclude that certain tetrapyrroles specifically inhibit the conversion of PrP-sen to PrP-res without apparent cytotoxic effects. The inhibition observed in the cell-free conversion reaction suggests that the mechanism involved direct interactions of the tetrapyrrole with PrP-res and/or PrP-sen. These findings introduce a new class of inhibitors of PrP-res formation that represents a potential source of therapeutic agents for transmissible spongiform encephalopathies.
Figures

References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
