

Nuclear-encoded proteins target to the plastid in Toxoplasma gondii and Plasmodium falciparum
- PMID: 9770490
- PMCID: PMC22835
- DOI: 10.1073/pnas.95.21.12352
Nuclear-encoded proteins target to the plastid in Toxoplasma gondii and Plasmodium falciparum
Abstract
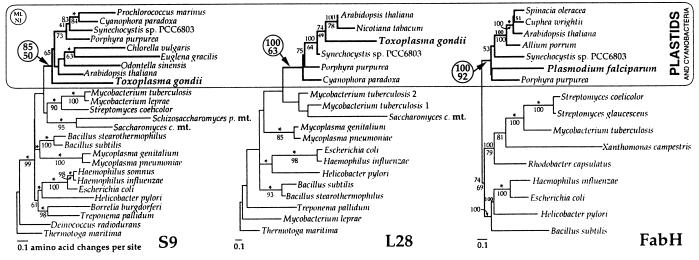
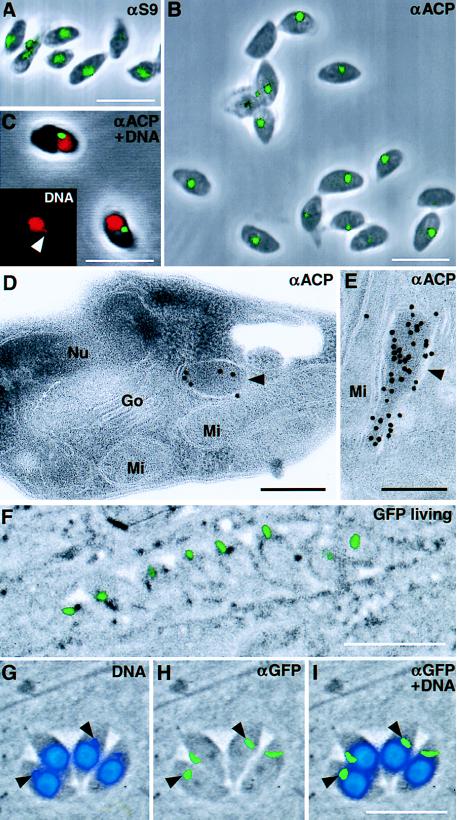
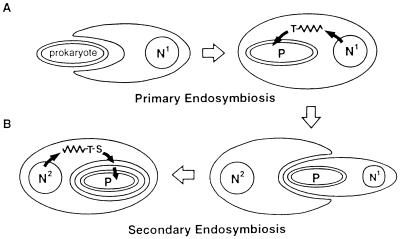
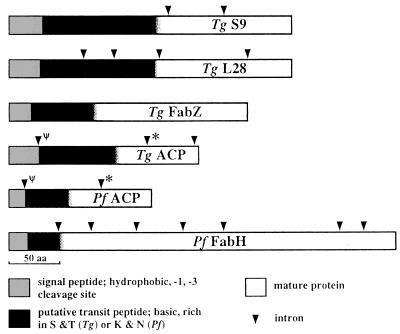
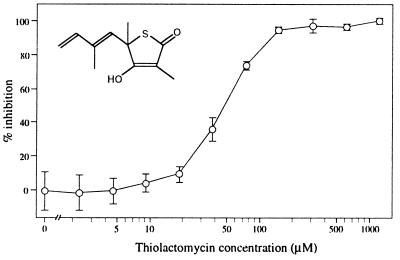
A vestigial, nonphotosynthetic plastid has been identified recently in protozoan parasites of the phylum Apicomplexa. The apicomplexan plastid, or "apicoplast," is indispensable, but the complete sequence of both the Plasmodium falciparum and Toxoplasma gondii apicoplast genomes has offered no clue as to what essential metabolic function(s) this organelle might perform in parasites. To investigate possible functions of the apicoplast, we sought to identify nuclear-encoded genes whose products are targeted to the apicoplast in Plasmodium and Toxoplasma. We describe here nuclear genes encoding ribosomal proteins S9 and L28 and the fatty acid biosynthetic enzymes acyl carrier protein (ACP), beta-ketoacyl-ACP synthase III (FabH), and beta-hydroxyacyl-ACP dehydratase (FabZ). These genes show high similarity to plastid homologues, and immunolocalization of S9 and ACP verifies that the proteins accumulate in the plastid. All the putatively apicoplast-targeted proteins bear N-terminal presequences consistent with plastid targeting, and the ACP presequence is shown to be sufficient to target a recombinant green fluorescent protein reporter to the apicoplast in transgenic T. gondii. Localization of ACP, and very probably FabH and FabZ, in the apicoplast implicates fatty acid biosynthesis as a likely function of the apicoplast. Moreover, inhibition of P. falciparum growth by thiolactomycin, an inhibitor of FabH, indicates a vital role for apicoplast fatty acid biosynthesis. Because the fatty acid biosynthesis genes identified here are of a plastid/bacterial type, and distinct from those of the equivalent pathway in animals, fatty acid biosynthesis is potentially an excellent target for therapeutics directed against malaria, toxoplasmosis, and other apicomplexan-mediated diseases.
Figures

References
-
- Wilson R J M, Denny P W, Preiser P R, Rangachari K, Roberts K, Roy A, Whyte A, Strath M, Moore D J, Moore P W, et al. J Mol Biol. 1996;261:155–172. - PubMed
-
- McFadden G I, Reith M, Munholland J, Lang-Unnasch N. Nature (London) 1996;381:482. - PubMed
-
- Köhler S, Delwiche C F, Denny P W, Tilney L G, Webster P, Wilson R J M, Palmer J D, Roos D S. Science. 1997;275:1485–1489. - PubMed
-
- Denny P, Preisser P, Williamson D, Wilson I. Protist. 1998;149:51–59. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
