

Selective regulation of apoptosis: the cytotoxic lymphocyte serpin proteinase inhibitor 9 protects against granzyme B-mediated apoptosis without perturbing the Fas cell death pathway
- PMID: 9774654
- PMCID: PMC109224
- DOI: 10.1128/MCB.18.11.6387
Selective regulation of apoptosis: the cytotoxic lymphocyte serpin proteinase inhibitor 9 protects against granzyme B-mediated apoptosis without perturbing the Fas cell death pathway
Abstract
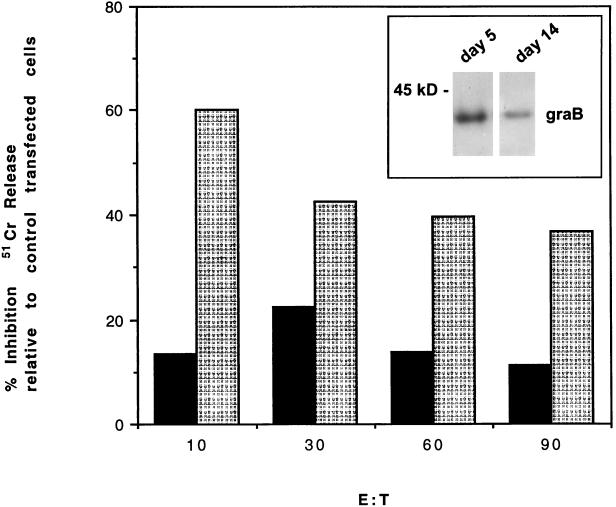
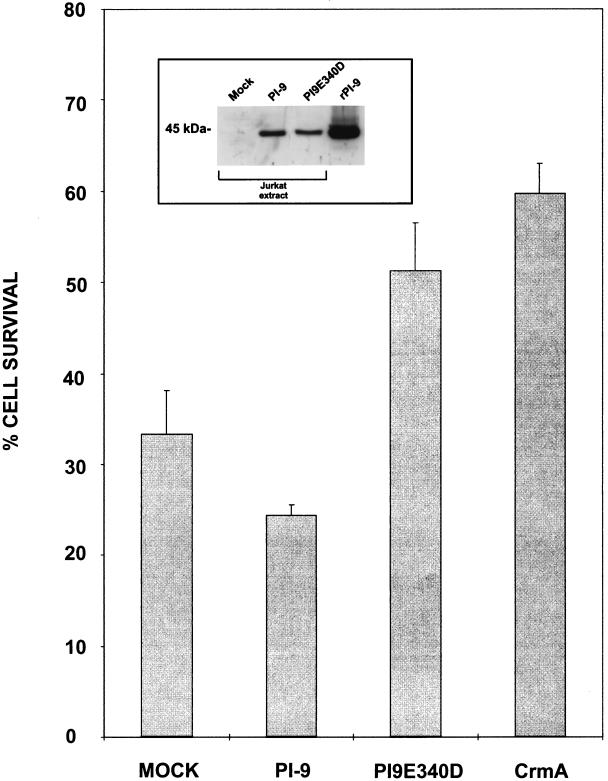
Cytotoxic lymphocytes (CLs) induce caspase activation and apoptosis of target cells either through Fas activation or through release of granule cytotoxins, particularly granzyme B. CLs themselves resist granule-mediated apoptosis but are eventually cleared via Fas-mediated apoptosis. Here we show that the CL cytoplasmic serpin proteinase inhibitor 9 (PI-9) can protect transfected cells against apoptosis induced by either purified granzyme B and perforin or intact CLs. A PI-9 P1 mutant (Glu to Asp) is a 100-fold-less-efficient granzyme B inhibitor that no longer protects against granzyme B-mediated apoptosis. PI-9 is highly specific for granzyme B because it does not inhibit eight of the nine caspases tested or protect transfected cells against Fas-mediated apoptosis. In contrast, the P1(Asp) mutant is an effective caspase inhibitor that protects against Fas-mediated apoptosis. We propose that PI-9 shields CLs specifically against misdirected granzyme B to prevent autolysis or fratricide, but it does not interfere with homeostatic deletion via Fas-mediated apoptosis.
Figures

Similar articles
-
A cytosolic granzyme B inhibitor related to the viral apoptotic regulator cytokine response modifier A is present in cytotoxic lymphocytes.J Biol Chem. 1996 Nov 1;271(44):27802-9. doi: 10.1074/jbc.271.44.27802. J Biol Chem. 1996. PMID: 8910377
-
Importance of the P4' residue in human granzyme B inhibitors and substrates revealed by scanning mutagenesis of the proteinase inhibitor 9 reactive center loop.J Biol Chem. 2001 May 4;276(18):15177-84. doi: 10.1074/jbc.M006645200. Epub 2001 Feb 1. J Biol Chem. 2001. PMID: 11278311
-
Granzyme B short-circuits the need for caspase 8 activity during granule-mediated cytotoxic T-lymphocyte killing by directly cleaving Bid.Mol Cell Biol. 2000 Jun;20(11):3781-94. doi: 10.1128/MCB.20.11.3781-3794.2000. Mol Cell Biol. 2000. PMID: 10805722 Free PMC article.
-
Serpins and regulation of cell death.Results Probl Cell Differ. 1998;24:63-89. doi: 10.1007/978-3-540-69185-3_4. Results Probl Cell Differ. 1998. PMID: 9949832 Review.
-
On the role and significance of Fas (Apo-1/CD95) ligand (FasL) expression in immune privileged tissues and cancer cells using multiple myeloma as a model.Leuk Lymphoma. 1998 Nov;31(5-6):477-90. doi: 10.3109/10428199809057607. Leuk Lymphoma. 1998. PMID: 9922038 Review.
Cited by
-
Granzyme B triggers a prolonged pressure to die in Bcl-2 overexpressing cells, defining a window of opportunity for effective treatment with ABT-737.Cell Death Dis. 2012 Jul 5;3(7):e344. doi: 10.1038/cddis.2012.73. Cell Death Dis. 2012. PMID: 22764103 Free PMC article.
-
Blockade of the granzyme B/perforin pathway through overexpression of the serine protease inhibitor PI-9/SPI-6 constitutes a mechanism for immune escape by tumors.Proc Natl Acad Sci U S A. 2001 Sep 25;98(20):11515-20. doi: 10.1073/pnas.201398198. Epub 2001 Sep 18. Proc Natl Acad Sci U S A. 2001. PMID: 11562487 Free PMC article.
-
The major human and mouse granzymes are structurally and functionally divergent.J Cell Biol. 2006 Nov 20;175(4):619-30. doi: 10.1083/jcb.200606073. J Cell Biol. 2006. PMID: 17116752 Free PMC article.
-
SerpinB1 regulates homeostatic expansion of IL-17+ γδ and CD4+ Th17 cells.J Leukoc Biol. 2014 Mar;95(3):521-30. doi: 10.1189/jlb.0613331. Epub 2013 Nov 18. J Leukoc Biol. 2014. PMID: 24249741 Free PMC article.
-
A functional genomics screen identifies PCAF and ADA3 as regulators of human granzyme B-mediated apoptosis and Bid cleavage.Cell Death Differ. 2014 May;21(5):748-60. doi: 10.1038/cdd.2013.203. Epub 2014 Jan 24. Cell Death Differ. 2014. PMID: 24464226 Free PMC article.
References
-
- Andersson S, Davis D L, Dahlback H, Jornvall H, Russell D W. Cloning, structure, and expression of the mitochondrial cytochrome P-450 sterol 26-hydroxylase, a bile acid biosynthetic enzyme. J Biol Chem. 1989;264:8222–8229. - PubMed
-
- Beatty K, Bieth J, Travis J. Kinetics of association of serine proteinases with native and oxidized alpha-1-proteinase inhibitor and alpha-1-antichymotrypsin. J Biol Chem. 1980;255:3931–3934. - PubMed
-
- Bump N J, Hackett M, Hugunin M, Seshagiri S, Brady K, Chen P, Ferenz C, Franklin S, Ghayur T, Li P, Licari P, Mankovich J, Shi L, Greenberg A H, Miller L K, Wong W W. Inhibition of ICE family proteases by baculovirus antiapoptotic protein p35. Science. 1995;269:1885–1888. - PubMed
-
- Cahill M A, Peter M E, Kischkel F C, Chinnaiyan A M, Dixit V M, Krammer P H, Nordheim A. CD95(APO-1/Fas) induces activation of SAP kinases downstream of ICE-like proteases. Oncogene. 1996;13:2087–2096. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous