

Targeted inactivation of mouse RAD52 reduces homologous recombination but not resistance to ionizing radiation
- PMID: 9774658
- PMCID: PMC109228
- DOI: 10.1128/MCB.18.11.6423
Targeted inactivation of mouse RAD52 reduces homologous recombination but not resistance to ionizing radiation
Abstract
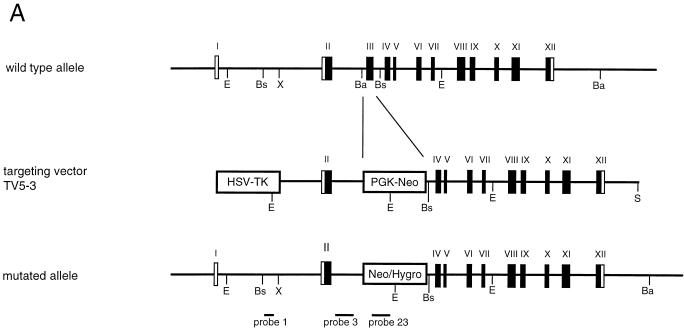
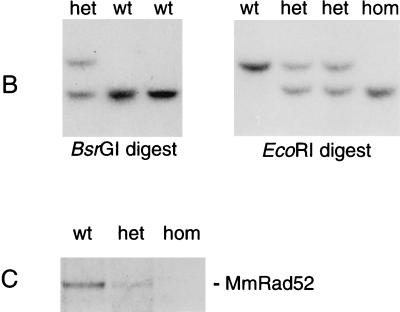
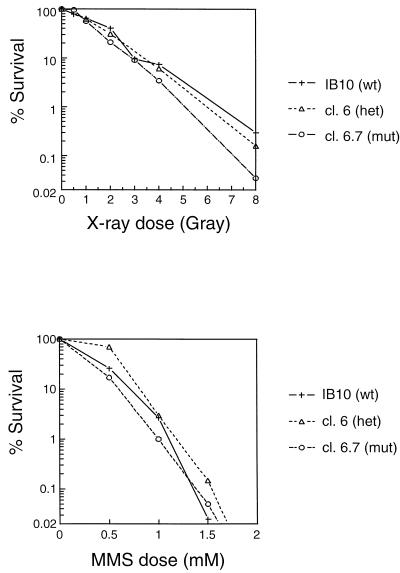
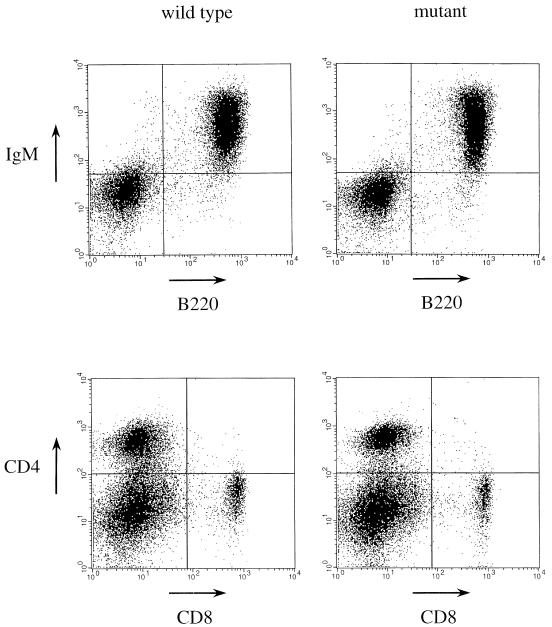
The RAD52 epistasis group is required for recombinational repair of double-strand breaks (DSBs) and shows strong evolutionary conservation. In Saccharomyces cerevisiae, RAD52 is one of the key members in this pathway. Strains with mutations in this gene show strong hypersensitivity to DNA-damaging agents and defects in recombination. Inactivation of the mouse homologue of RAD52 in embryonic stem (ES) cells resulted in a reduced frequency of homologous recombination. Unlike the yeast Scrad52 mutant, MmRAD52(-/-) ES cells were not hypersensitive to agents that induce DSBs. MmRAD52 null mutant mice showed no abnormalities in viability, fertility, and the immune system. These results show that, as in S. cerevisiae, MmRAD52 is involved in recombination, although the repair of DNA damage is not affected upon inactivation, indicating that MmRAD52 may be involved in certain types of DSB repair processes and not in others. The effect of inactivating MmRAD52 suggests the presence of genes functionally related to MmRAD52, which can partly compensate for the absence of MmRad52 protein.
Figures
References
-
- Akaboshi E, Inoue Y, Ryo H. Cloning of the cDNA and genomic DNA that correspond to the recA-like gene of Drosophila melanogaster. Jpn J Genet. 1994;69:663–670. - PubMed
-
- Bai Y, Symington L S. A Rad52 homolog is required for RAD51-independent mitotic recombination in Saccharomyces cerevisiae. Genes Dev. 1996;10:2025–2037. - PubMed
-
- Baumann P, Benson F E, West S C. Human Rad51 protein promotes ATP-dependent homologous pairing and strand transfer reactions in vitro. Cell. 1996;87:757–766. - PubMed
-
- Bendixen C, Sunjevaric I, Bauchwitz R, Rothstein R. Identification of a mouse homologue of the Saccharomyces cerevisiae recombination and repair gene, RAD52. Genomics. 1994;23:300–303. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials