

Novel cofactors and TFIIA mediate functional core promoter selectivity by the human TAFII150-containing TFIID complex
- PMID: 9774672
- PMCID: PMC109242
- DOI: 10.1128/MCB.18.11.6571
Novel cofactors and TFIIA mediate functional core promoter selectivity by the human TAFII150-containing TFIID complex
Abstract
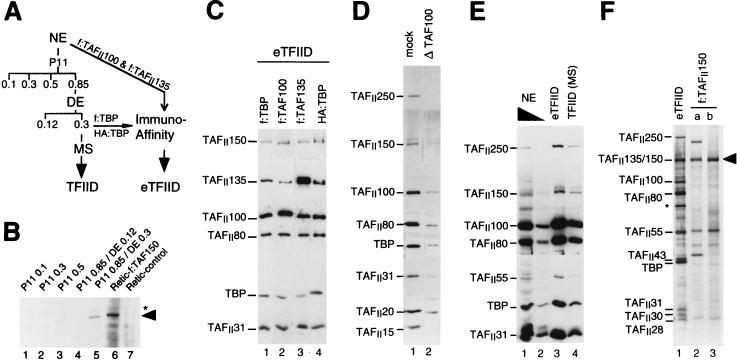
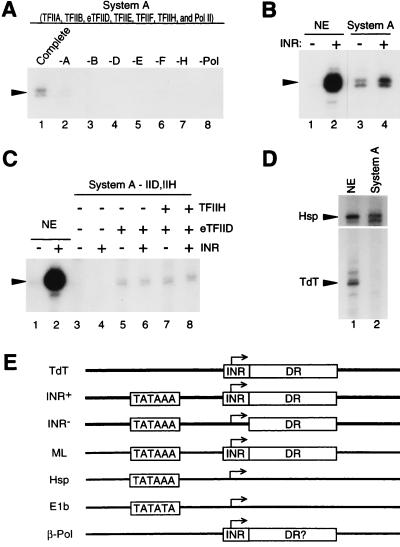
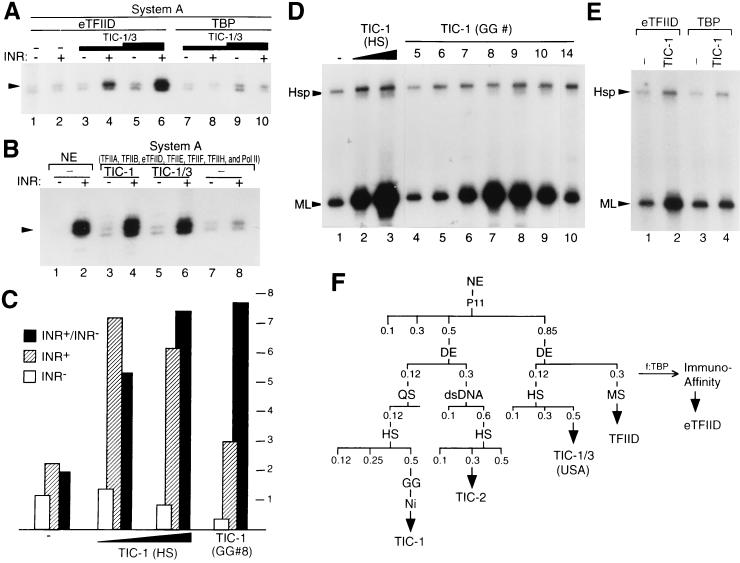
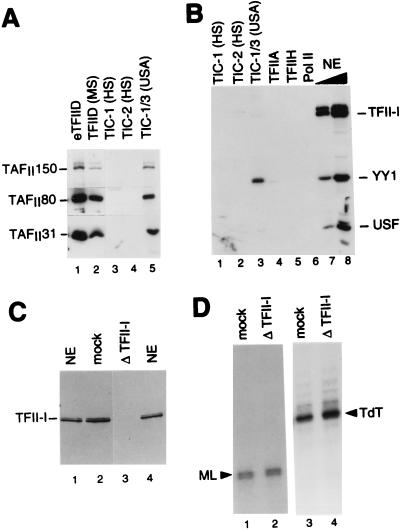
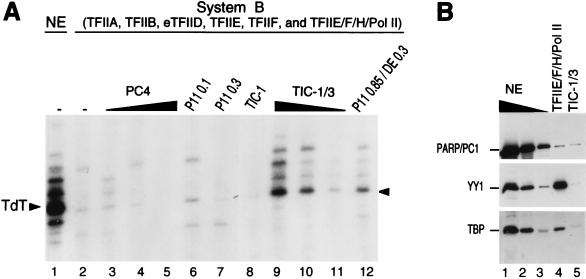
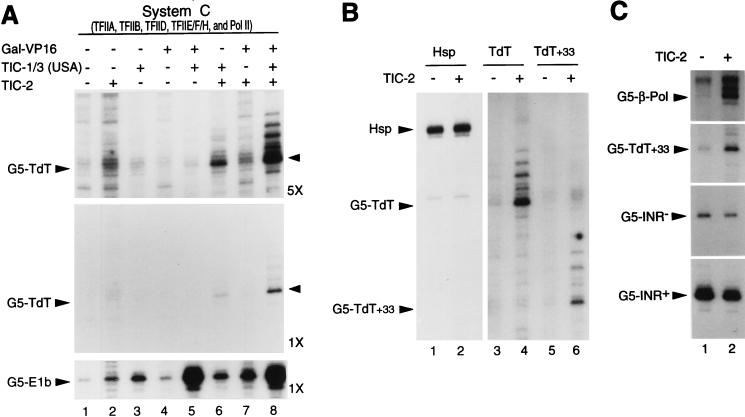
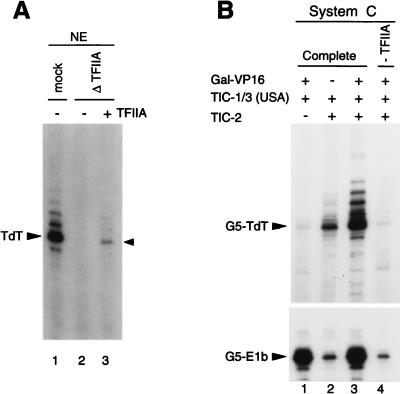
TATA-binding protein-associated factors (TAFIIs) within TFIID control differential gene transcription through interactions with both activators and core promoter elements. In particular, TAFII150 contributes to initiator-dependent transcription through an unknown mechanism. Here, we address whether TAFIIs within TFIID are sufficient, in conjunction with highly purified general transcription factors (GTFs), for differential core promoter-dependent transcription by RNA polymerase II and whether additional cofactors are required. We identify the human homologue of Drosophila TAFII150 through cognate cDNA cloning and show that it is a tightly associated component of human TFIID. More importantly, we demonstrate that the human TAFII150-containing TFIID complex is not sufficient, in the context of all purified GTFs and RNA polymerase II, to mediate transcription synergism between TATA and initiator elements and initiator-directed transcription from a TAFII-dependent TATA-less promoter. Therefore, TAFII-promoter interactions are not sufficient for the productive core promoter-selective functions of TFIID. Consistent with this finding, we have partially purified novel cofactor activities (TICs) that potentiate the TAFII-mediated synergism between TATA and initiator elements (TIC-1) and TAFII-dependent transcription from TATA-less promoters (TIC-2 and -3). Furthermore, we demonstrate an essential function for TFIIA in TIC- and TAFII-dependent basal transcription from a TATA-less promoter. Our results reveal a parallel between the basal transcription activity of TAFIIs through core promoter elements and TAFII-dependent activator function.
Figures

Similar articles
-
Taspase1 processing alters TFIIA cofactor properties in the regulation of TFIID.Transcription. 2015;6(2):21-32. doi: 10.1080/21541264.2015.1052178. Transcription. 2015. PMID: 25996597 Free PMC article.
-
A transcription factor IIA-binding site differentially regulates RNA polymerase II-mediated transcription in a promoter context-dependent manner.J Biol Chem. 2017 Jul 14;292(28):11873-11885. doi: 10.1074/jbc.M116.770412. Epub 2017 May 24. J Biol Chem. 2017. PMID: 28539359 Free PMC article.
-
Transcription factor IIA derepresses TATA-binding protein (TBP)-associated factor inhibition of TBP-DNA binding.J Biol Chem. 1998 Jun 5;273(23):14293-300. doi: 10.1074/jbc.273.23.14293. J Biol Chem. 1998. PMID: 9603936
-
The general transcription machinery and general cofactors.Crit Rev Biochem Mol Biol. 2006 May-Jun;41(3):105-78. doi: 10.1080/10409230600648736. Crit Rev Biochem Mol Biol. 2006. PMID: 16858867 Review.
-
TAF(II)250: a transcription toolbox.J Cell Sci. 2001 Aug;114(Pt 16):2895-902. doi: 10.1242/jcs.114.16.2895. J Cell Sci. 2001. PMID: 11686293 Review.
Cited by
-
Taspase1 processing alters TFIIA cofactor properties in the regulation of TFIID.Transcription. 2015;6(2):21-32. doi: 10.1080/21541264.2015.1052178. Transcription. 2015. PMID: 25996597 Free PMC article.
-
The general transcription factors IIA, IIB, IIF, and IIE are required for RNA polymerase II transcription from the human U1 small nuclear RNA promoter.Mol Cell Biol. 1999 Mar;19(3):2130-41. doi: 10.1128/MCB.19.3.2130. Mol Cell Biol. 1999. PMID: 10022900 Free PMC article.
-
The YEATS domain of Taf14 in Saccharomyces cerevisiae has a negative impact on cell growth.Mol Genet Genomics. 2010 Apr;283(4):365-80. doi: 10.1007/s00438-010-0523-x. Epub 2010 Feb 24. Mol Genet Genomics. 2010. PMID: 20179968 Free PMC article.
-
Prevalence of the initiator over the TATA box in human and yeast genes and identification of DNA motifs enriched in human TATA-less core promoters.Gene. 2007 Mar 1;389(1):52-65. doi: 10.1016/j.gene.2006.09.029. Epub 2006 Oct 10. Gene. 2007. PMID: 17123746 Free PMC article.
-
Core promoter-specific gene regulation: TATA box selectivity and Initiator-dependent bi-directionality of serum response factor-activated transcription.Biochim Biophys Acta. 2016 Apr;1859(4):553-63. doi: 10.1016/j.bbagrm.2016.01.005. Epub 2016 Jan 26. Biochim Biophys Acta. 2016. PMID: 26824723 Free PMC article.
References
-
- Burley S K, Roeder R G. Biochemistry and structural biology of transcription factor IID (TFIID) Annu Rev Biochem. 1996;65:769–799. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases