

doi: 10.1128/MCB.18.11.6826.
Nuclear mRNA export requires complex formation between Mex67p and Mtr2p at the nuclear pores
Affiliations
- PMID: 9774696
- PMCID: PMC109266
- DOI: 10.1128/MCB.18.11.6826
Item in Clipboard
Nuclear mRNA export requires complex formation between Mex67p and Mtr2p at the nuclear pores
Mol Cell Biol.
1998 Nov.
Abstract
We have identified between Mex67p and Mtr2p a complex which is essential for mRNA export. This complex, either isolated from yeast or assembled in Escherichia coli, can bind in vitro to RNA through Mex67p. In vivo, Mex67p requires Mtr2p for association with the nuclear pores, which can be abolished by mutating either MEX67 or MTR2. In all cases, detachment of Mex67p from the pores into the cytoplasm correlates with a strong inhibition of mRNA export. At the nuclear pores, Nup85p represents one of the targets with which the Mex67p-Mtr2p complex interacts. Thus, Mex67p and Mtr2p constitute a novel mRNA export complex which can bind to RNA via Mex67p and which interacts with nuclear pores via Mtr2p.
Figures
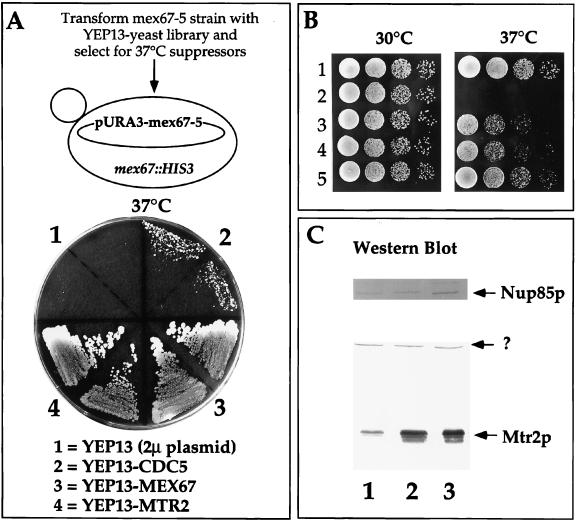
Genetic interaction between MEX67 and MTR2. (A) Isolation of the MTR2 gene as a high-copy-number suppressor of thermosensitive mex67-5. A YPD plate contained the thermosensitive mex67-5 strain transformed with the indicated high-copy-number 2μm (2μ) plasmids and grown for 3 days at 37°C. Note that YEP13-CDC5 does not complement the growth defect of thermosensitive mex67-5 as well as does YEP13-MTR2 at 37°C. (B) Growth of MEX67+ and mex67-5 cells transformed with various plasmids at 30 and 37°C. Rows: 1, haploid RS453; 2 to 5, thermosensitive mex67-5 cells transformed with low-copy-number plasmids pRS316 (2) and pRS316-MTR2 (3) and high-copy-number plasmids YEP13-MTR2 (4) and pRS316-MEX67 (5). (C) Expression of Mtr2p in mex67-5 yeast strains. Whole-cell SDS lysates from mex67-5 cells transformed with low-copy-number plasmids pRS316 (lane 1) and pRS316-MTR2 (lane 2) and high-copy-number plasmid YEP13-MTR2 (lane 3) were analyzed by Western blotting with anti-Mtr2p and anti-Nup85p antibodies. The band marked by a question mark was cross-reactive with anti-Mtr2p antibodies.
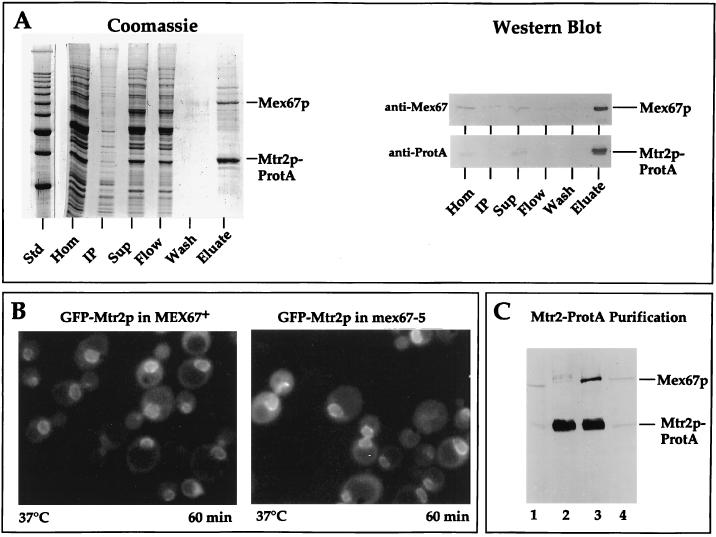
Mtr2p and Mex67p form a complex at nuclear pores. (A) Affinity purification of Mtr2p-ProtA by IgG-Sepharose chromatography. A cell homogenate (Hom), an insoluble pellet (IP), a soluble supernatant (Sup), the flowthrough (Flow), a pH 5 wash fraction (Wash), and a 300-fold equivalent of the eluted complex (Eluate) were analyzed by SDS-PAGE followed by Coomassie blue staining or by Western blotting with IgG coupled to horseradish peroxidase to detect the ProtA moiety (anti-ProtA) or with anti-Mex67p antibodies (anti-Mex67). The positions of Mex67p and Mtr2p-ProtA are shown. Std, marker proteins (10-kDa ladder with a stronger 50-kDa band). (B) Nuclear envelope location of GFP-Mtr2p in MEX67+ and thermosensitive mex67-5 cells as revealed by fluorescence microscopy. MEX67/GFP-MTR2 and mex67-5/GFP-MTR2 cells (Table 1) were shifted for 60 min to 37°C before pictures were taken. Under the same conditions, thermosensitive cells expressing Mex67-5–GFP showed cytoplasmic mislocalization (40). (C) Affinity purification of Mtr2p-ProtA from thermosensitive mex67-5 (lanes 1 and 2) and MEX67+ (lanes 3 and 4) cells grown at 30°C by IgG-Sepharose chromatography. A whole-cell lysate (lanes 1 and 4) and the eluate fraction from the IgG-Sepharose column (lanes 2 and 3) were analyzed by SDS-PAGE and Western blotting with antibodies against Mtr2p-ProtA and Mex67p.
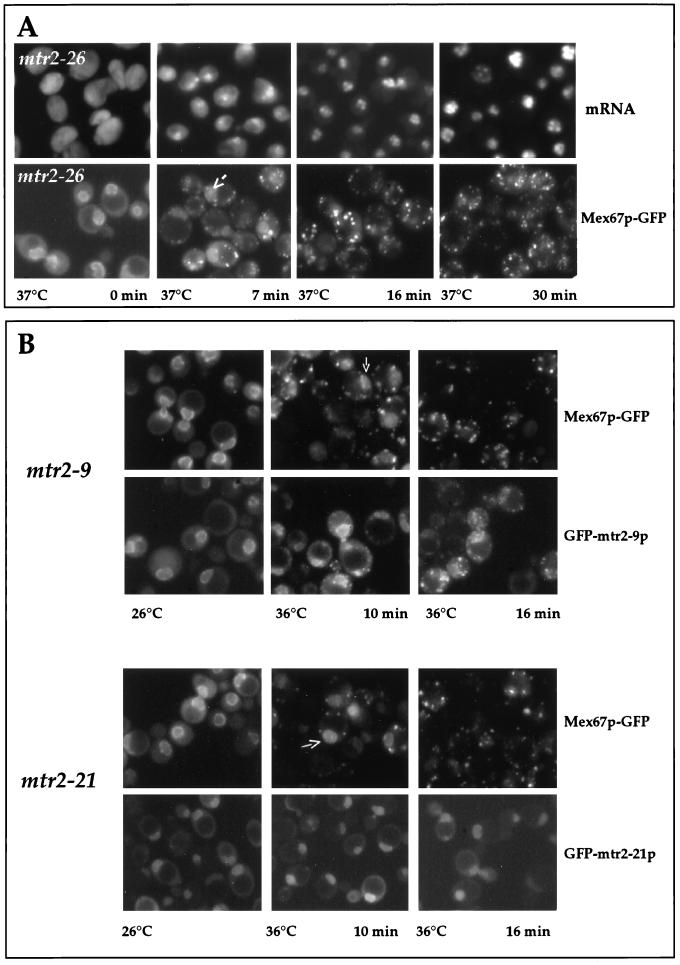
Nuclear mRNA export and localization of GFP-tagged Mtr2p and Mex67p in mtr2 mutant cells. (A) Correlation between inhibition of mRNA export and cytoplasmic mislocalization of Mex67p-GFP in mtr2-26 cells. mtr2-26 cells were grown at 26°C before a shift to 37°C. Samples were taken at the indicated times and analyzed for mRNA export defects [by in situ hybridization with a Cy3-labeled oligo(dT) probe] and for Mex67p-GFP mislocalization. (B) Localization of Mex67p-GFP and GFP-Mtr2p mutant proteins in living cells. mtr2-9/MEX67-GFP, GFP-mtr2-9, mtr2-21/MEX67-GFP, and GFP-mtr2-21 cells (Table 1) were grown at 26°C before a shift to 36°C for 0, 10, and 16 min. The corresponding GFP-tagged proteins were localized in the cells by fluorescence microscopy.
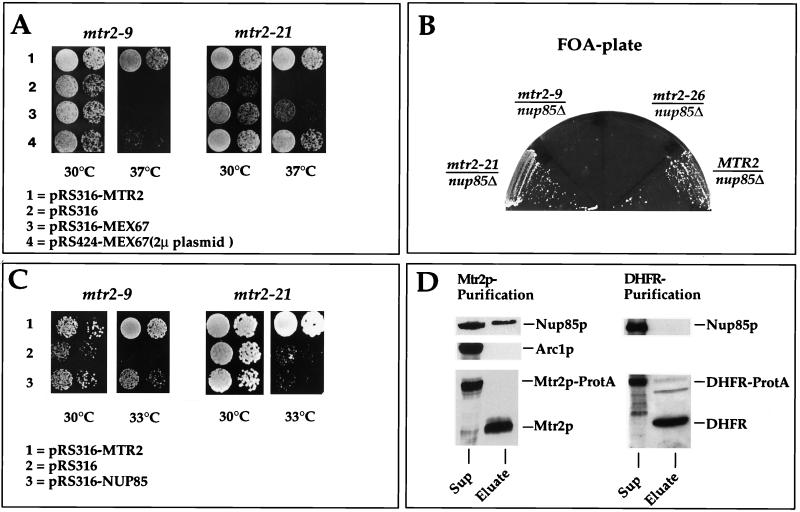
Gentic and physical interactions between Mtr2p and Nup85p. (A) Suppression of the thermosensitive mtr2-9 and mtr2-21 growth defect at 37°C by overexpression of MEX67. Precultures were diluted in growth medium, and equivalent amounts of cells (diluted in 10−1 steps) were spotted onto YPD plates. Plates were incubated for 3 days. (B) Genetic interaction between thermosensitive nup85Δ and mtr2 alleles. Strain nup85Δ/MTR2 shuffle (Table 1) was transformed with plasmids containing the MTR2, mtr2-26, mtr2-9, and mtr2-21 genes. The synthetic lethal relationship between the nup85Δ and mtr2 alleles was tested by streaking transformants on FOA-containing plates. Growth inhibition on FOA plates indicates synthetic lethality. (C) Complementation of the thermosensitive mtr2-9 and mtr2-21 growth defect at 33°C by overexpression of NUP85. Precultures were diluted in growth medium, and equivalent amounts of cells (diluted in 10−1 steps) were spotted onto YPD plates. Plates were incubated for 3 days. (D) Western blot analysis of purified Mtr2p reveals an association with Nup85p. Mtr2p-TEV-ProtA and Prot-TEV-DHFR were affinity purified as described in Materials and Methods. Each TEV eluate (25 μl) was analyzed by SDS-PAGE followed by Western blotting with anti-Mtr2p, anti-Nup85p, anti-Arc1p, and anti-DHFR antibodies. The blots were developed with the Amersham ECL kit. Sup, supernatant.
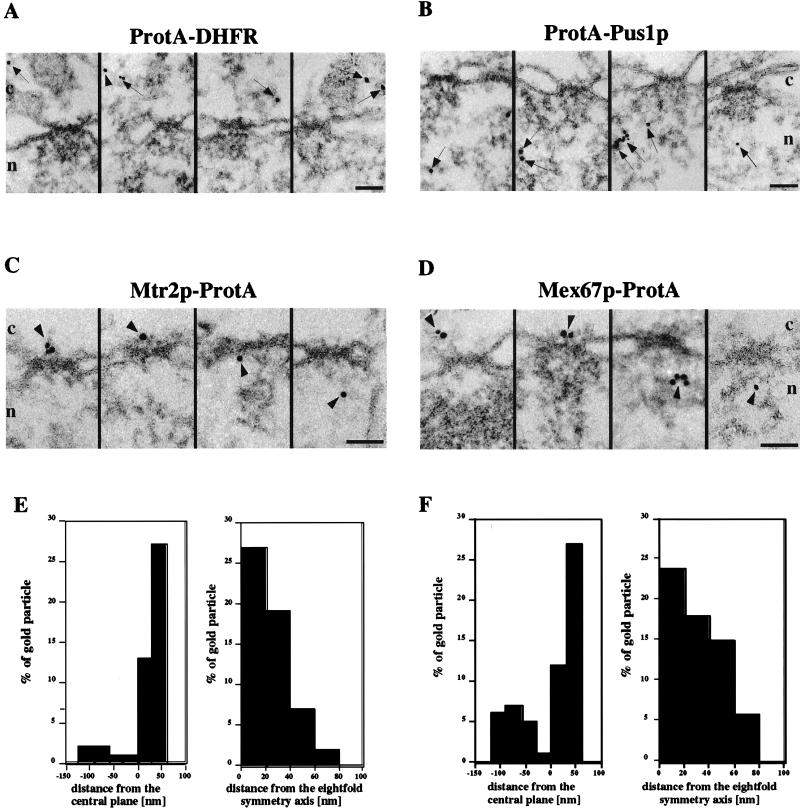
Immunoelectron microscopy of Mex67p and Mtr2p. Nuclear envelope cross sections with adjacent nucleoplasm and cytoplasm from Triton X-100-extracted cells expressing ProtA-DHFR (A), ProtA-Pus1p (B), Mtr2p-ProtA (C), and Mex67p-ProtA (D). Cells were preembedding labeled with gold-conjugated anti-ProtA antibody. A gallery of representative photographs is shown. For Mtr2p-ProtA and Mex67p-ProtA, selected examples of labeled NPCs and the quantitative analysis of gold particles associated with NPCs (E and F) are also shown. The distances of gold particles relative to the twofold symmetry axis of the NPC (axis parallel to the nuclear envelope passing through the center of the NPC, with positive distances representing the cytoplasmic side of the NPC) and the eightfold symmetry axis of the NPC (axis perpendicular to the nuclear envelope and passing through the center of the NPC) were determined in cross sections. Cross-sectioned nuclear envelopes from two experiments which yielded 38 cells and 65 gold particles associated with the NPC for Mex67p and 32 cells and 55 gold particles associated with the NPC for Mtr2p were analyzed. The arrows indicate gold particles in the cytoplasm and the nucleus, whereas the arrowheads indicate gold particles associated with NPCs. Cytoplasmic (c) and nuclear (n) sides of the nuclear envelope are indicated. Scale bars, 100 nm.
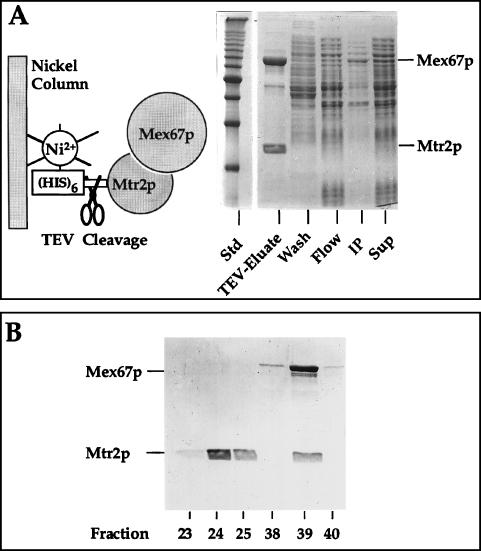
Purification of recombinant Mtr2p and Mex67p from E. coli. (A) Coexpression and complex formation of Mtr2p and Mex67p in bacteria. His6-TEV-Mtr2p and untagged Mex67p were expressed and purified from E. coli. A soluble supernatant (Sup), an insoluble pellet (IP), the flowthrough (Flow), a wash fraction (Wash), and the TEV protease-eluted complex (TEV-Eluate) were analyzed by SDS-PAGE and Coomassie blue staining. The positions of Mex67p and Mtr2p are shown. Std, marker proteins (10-kDa ladder). (B) Purification of the recombinant Mex67p-Mtr2p complex by ion-exchange chromatography. Fractions from the MonoS column (23 to 25 and 38 to 40) were analyzed by SDS-PAGE and Coomassie blue staining. The two weakly staining bands below full-length Mex67p were cross-reactive with anti-Mex67p antibodies and thus may correspond to proteolytic breakdown products. Note that Mtr2p tends to migrate as a broad and fuzzy band.
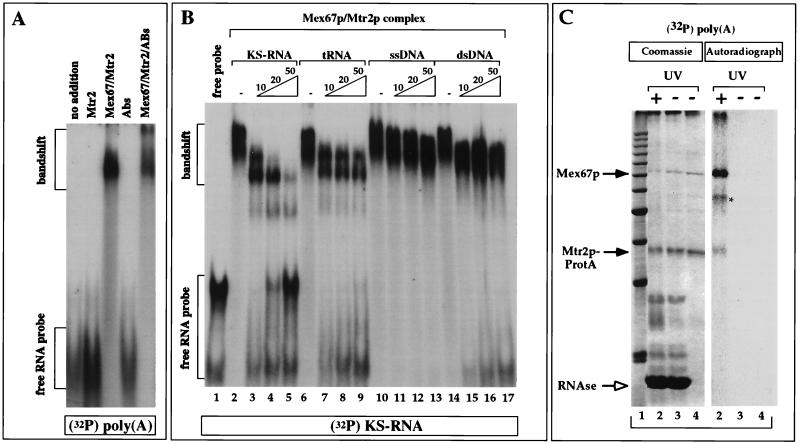
Band shift assays and UV cross-linking to detect the binding of Mex67p to RNA. (A) Binding to RNA was tested in a band shift assay with in vitro-transcribed, 32P-radiolabeled poly(A)+ RNA and purified recombinant proteins in the absence or presence of anti-Mex67p antibodies (Abs or ABs). (B) Competition of the formation of the 32P-labeled KS-RNA–Mex67p–Mtr2p complex by an excess (10-, 20-, or 50-fold) of unlabeled KS-RNA, tRNA, single-stranded DNA (ssDNA), and double-stranded DNA (dsDNA). 32P-labeled KS-RNA (5 ng) was incubated with 1 μg of E. coli-derived, purified Mex67p-Mtr2p complex in the absence (−) or presence of increasing amounts of cold competitor KS-RNA, yeast tRNAMet, single-stranded DNA, and double-stranded DNA. (C) In vitro UV cross-linking of Mex67p to RNA. The Mex67p-Mtr2p-ProtA complex bound to IgG-Sepharose beads was incubated with 1 ng of 32P-labeled poly(A)+ RNA and UV irradiated before analysis by SDS-PAGE and Coomassie blue staining or autoradiography. Lane 1, 10-kDa marker proteins; lanes 2, UV-irradiated beads treated with RNase; lanes 3, non-UV-irradiated beads treated with RNase; lanes 4, input of beads. Proteins were finally released from the beads with SDS. The positions of Mex67p and Mtr2p-ProtA are indicated by arrows. The asterisk indicates an unidentified band.
References
-
- Amberg D C, Goldstein A L, Cole C N. Isolation and characterization of RAT1: an essential gene of Saccharomyces cerevisiae required for the efficient nucleocytoplasmic trafficking of mRNA. Genes Dev. 1992;6:1173–1189. - PubMed
-
- Doye V, Hurt E C. From nucleoporins to nuclear pore complexes. Curr Opin Cell Biol. 1997;9:401–411. - PubMed
-
- Fahrenkrog, B., E. Hurt, U. Aebi, and N. Panté. Submitted for publication.
-
- Fischer U, Huber J, Boelens W C, Mattaj I W, Lührmann R. The HIV-1 Rev activation domain is a nuclear export signal that accesses an export pathway used by specific cellular RNAs. Cell. 1995;82:475–483. - PubMed
-
- Fornerod M, Ohno M, Yoshida M, Mattaj I W. CRM1 is an export receptor for leucine-rich nuclear export signals. Cell. 1997;90:1051–1060. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases