

Modulation of endothelial cell function by normal polyspecific human intravenous immunoglobulins: a possible mechanism of action in vascular diseases
- PMID: 9777957
- PMCID: PMC1853054
- DOI: 10.1016/S0002-9440(10)65670-2
Modulation of endothelial cell function by normal polyspecific human intravenous immunoglobulins: a possible mechanism of action in vascular diseases
Abstract
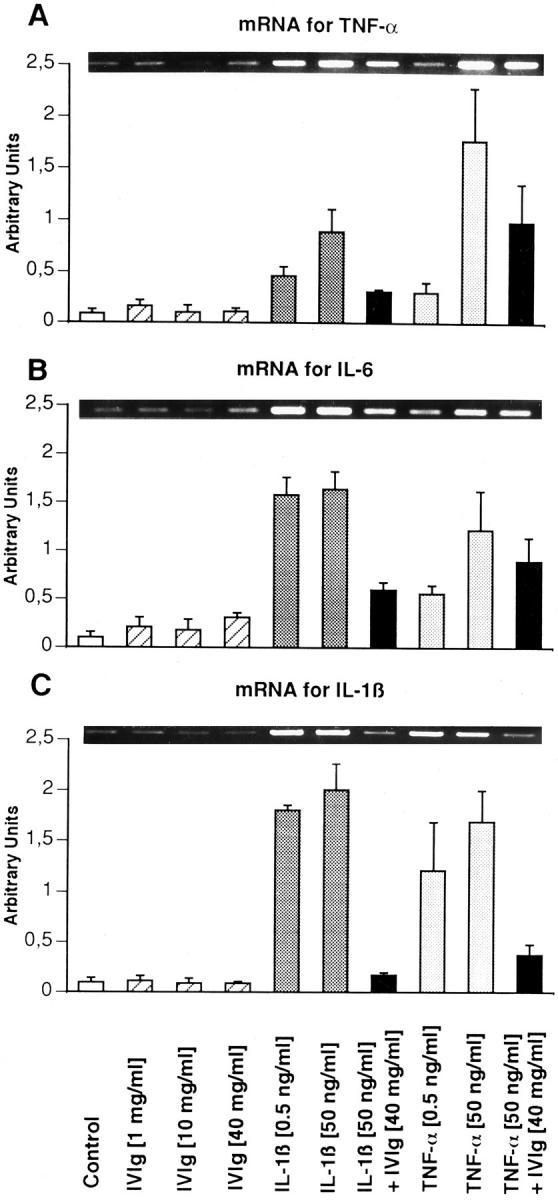
Intravenous immunoglobulin (IVIg) is increasingly used in the treatment of autoimmune and inflammatory diseases, including vasculitides and Kawasaki disease. However, the outcome of IVIg interaction with endothelial cells of the vascular bed is not clear as yet. We have investigated the effect of IVIg on the in vitro activation of human endothelial cells, as assessed by cell proliferation and reverse transcription-polymerase chain reaction-detected expression of mRNA coding for adhesion molecules (intercellular adhesion molecule-1 and vascular cellular adhesion molecule-1), chemokines (monocyte chemoattractant protein-1, macrophage colony-stimulating factor, and granulocyte-macrophage colony-stimulating factor), and proinflammatory cytokines (tumor necrosis factor-alpha, interleukin-1beta, and interleukin-6). IVIg inhibited proliferation of endothelial cells in a time-dependent manner. This effect was dependent on both Fc and F(ab')2 fragments of the immunoglobulin molecule and was fully reversible. Tumor necrosis factor-alpha and interleukin-1beta also inhibited thymidine incorporation, but to a lesser degree. IVIg had no effect on basal levels of mRNA coding for the adhesion molecules, chemokines, and proinflammatory cytokines. IVIg fully down-regulated the expression induced by tumor necrosis factor-alpha or interleukin-1beta of mRNA coding for these molecules. Thus, blockade of cellular proliferation and of cytokine-induced expression of adhesion molecules, chemokines, and cytokines may explain the therapeutic effect of IVIg in vascular and inflammatory disorders.
Figures








References
-
- Carlos TM, Harlan JM: Leukocyte-endothelial adhesion molecules. Blood 1994, 84:2068-2101 - PubMed
-
- Demuth K, Myara I, Moatti N: Biologie de la cellule endothéliale et athérogenèse. Ann Biol Clin 1995, 53:171-189 - PubMed
-
- Matsushima KV, Oppenheim JJ: Interleukin-8 and MCAF: novel inflammatory cytokines induced by TNF and Il-1. Cytokines 1989, 1:2-10 - PubMed
-
- Brady HR: Leukocyte adhesion molecules and kidney diseases. Kidney Int 1994, 45:1285-1300 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
