

Two types of parasympathetic preganglionic neurones in the superior salivatory nucleus characterized electrophysiologically in slice preparations of neonatal rats
- PMID: 9782167
- PMCID: PMC2231255
- DOI: 10.1111/j.1469-7793.1998.157by.x
Two types of parasympathetic preganglionic neurones in the superior salivatory nucleus characterized electrophysiologically in slice preparations of neonatal rats
Abstract
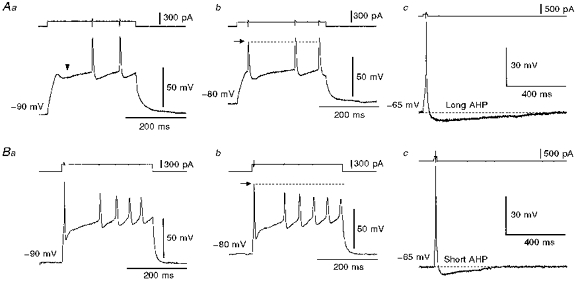
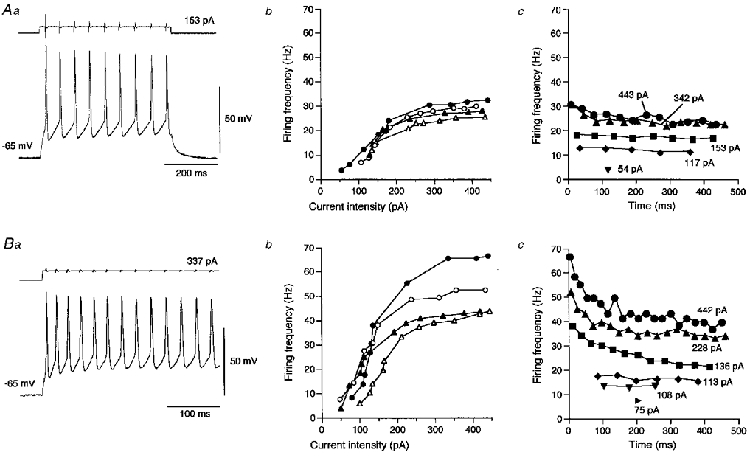
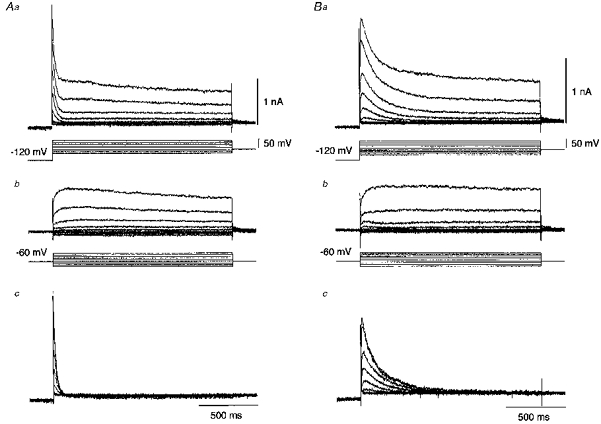
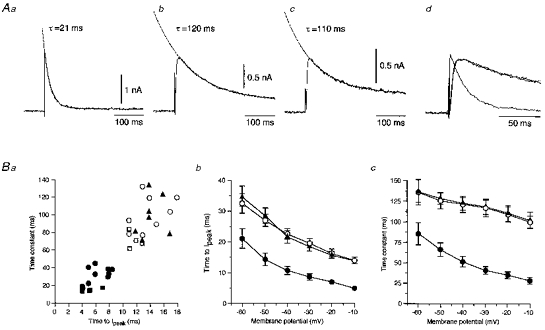
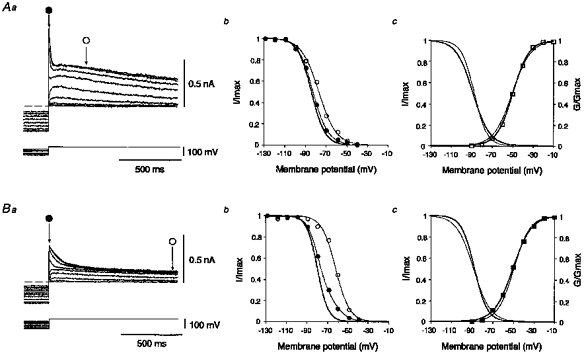
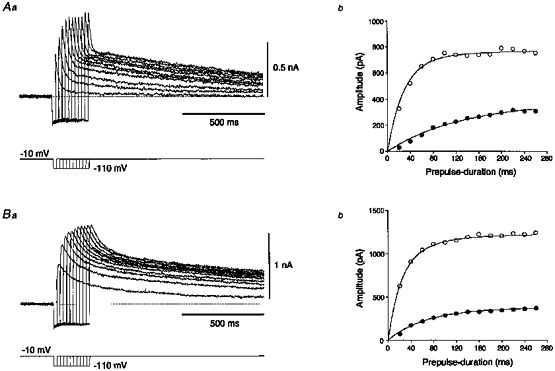
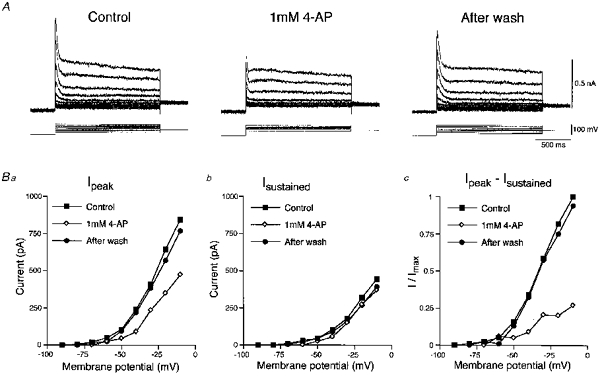
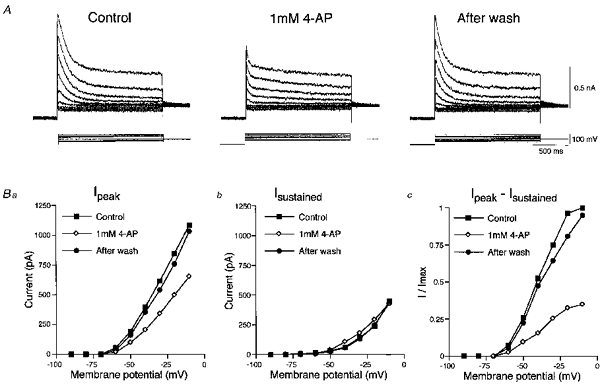
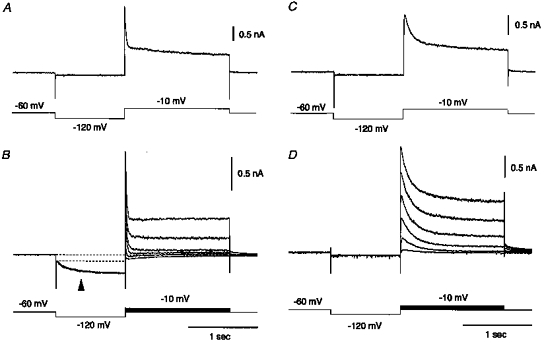
1. The electrophysiological properties of parasympathetic preganglionic neurones in the superior salivatory nucleus were studied in thin- and thick-slice preparations of rats aged 1 and 2 weeks using the whole-cell patch-clamp technique. 2. The superior salivatory neurones were identified by a retrograde tracing method with dextran-tetramethylrhodamine-lysine. The injection of the tracer into the chorda-lingual nerve labelled the neurones innervating the submandibular ganglia and those innervating the intra-lingual ganglia, while the injection into the tip of the tongue labelled the latter group of neurones. 3. Firing characteristics were investigated mainly in the neurones of 6-8 days postnatal rats. In response to an injection of long depolarizing current pulses at hyperpolarized membrane potentials (< -80 mV) under a current clamp, the neurones labelled from the nerve displayed a train of action potentials with either a long silent period preceding the first spike (late spiking pattern) or a long silent period interposed between the first and second spikes (interrupted spiking pattern). The neurones labelled from the tongue invariably displayed the interrupted spiking pattern. 4. Under a voltage clamp, among the neurones from 6-8 days postnatal rats, those labelled from the nerve expressed either a fast or a slow transient outward current (A-current), while those labelled from the tongue invariably showed a slow transient outward current. Both the fast and slow A-currents were largely depressed by 1 mM 4-aminopyridine. 5. Similar fast and slow A-currents were observed in the neurones of rats aged 14-15 days. Both the time to peak and decay time constant of these A-currents were accelerated, suggesting a developmental trend of maturation in the activation and inactivation kinetics between 6 and 15 days postnatal. 6. Based on the differences in the firing pattern and outward current, the superior salivatory neurones can be separated into two distinct types. We discuss the functional aspects of these two types of neurones with reference to their target organs.
Figures

References
-
- Bayliss DA, Viana F, Bellingham MC, Berger AJ. Characteristics and postnatal development of a hyperpolarization-activated inward current in rat hypoglossal motoneurons in vitro. Journal of Neurophysiology. 1994;71:119–128. - PubMed
-
- Bordey A, Feltz P, Trouslard J. Kinetics of A-currents in sympathetic preganglionic neurons and glial cells. NeuroReport. 1995;7:37–40. - PubMed
-
- Bottaro B, Cutler LS. An electrophysiological study of the postnatal development of the autonomic innervation of the rat submandibular salivary gland. Archives of Oral Biology. 1984;29:237–242. - PubMed
-
- Bylund DB, Martinez JR, Camden J, Jones SB. Autonomic receptors in the developing submandibular glands of neonatal rats. Archives of Oral Biology. 1982;27:945–950. - PubMed
-
- Carrier GO. Whole-cell and perforated patch recordings of four distinct K+ currents in acutely dispersed coeliac-superior mesenteric ganglia neurones of adult rats. Brain Research. 1995;701:1–12. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources