

Onset of keratin 17 expression coincides with the definition of major epithelial lineages during skin development
- PMID: 9786956
- PMCID: PMC2132846
- DOI: 10.1083/jcb.143.2.469
Onset of keratin 17 expression coincides with the definition of major epithelial lineages during skin development
Abstract
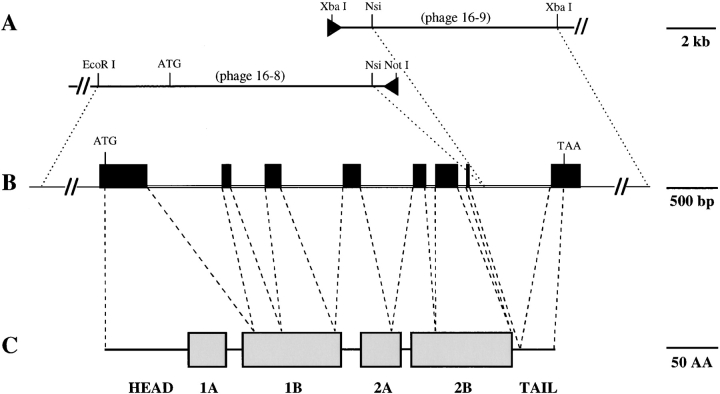
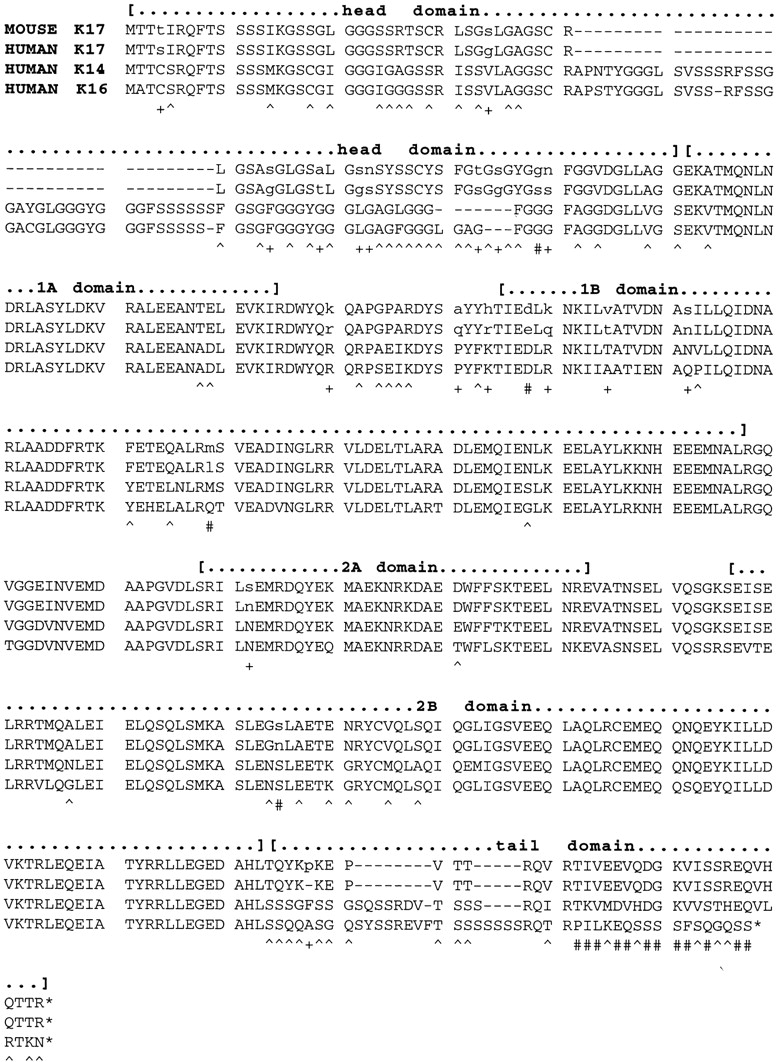
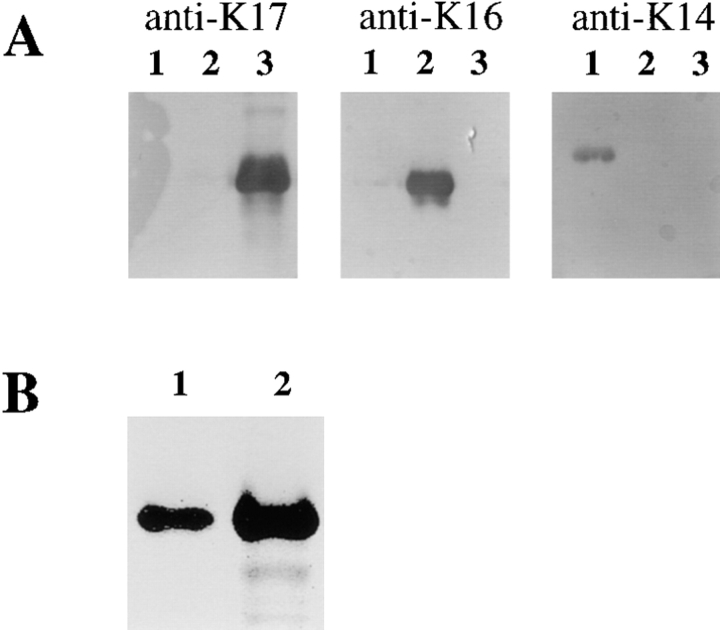
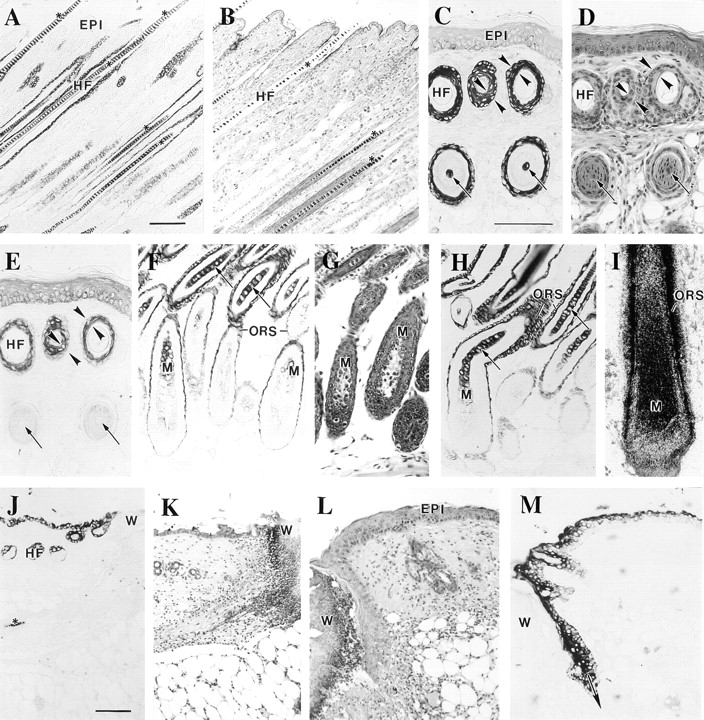
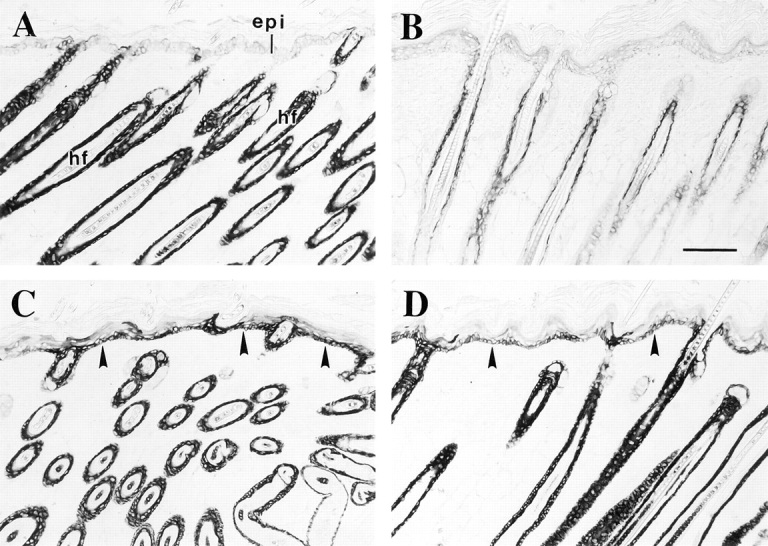
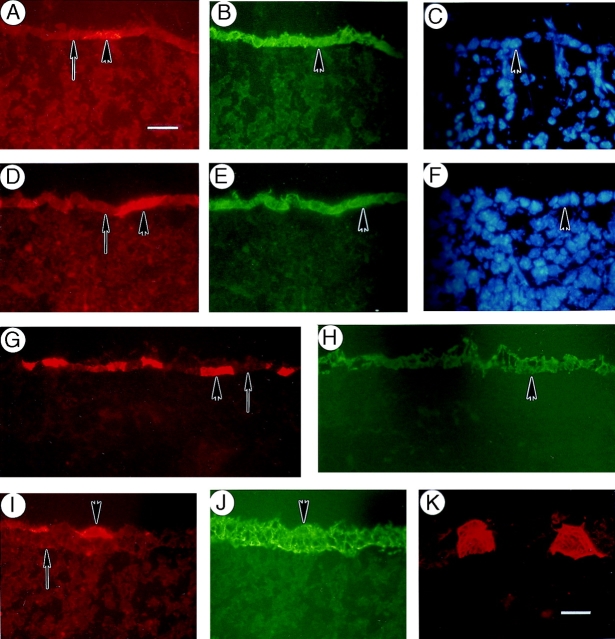
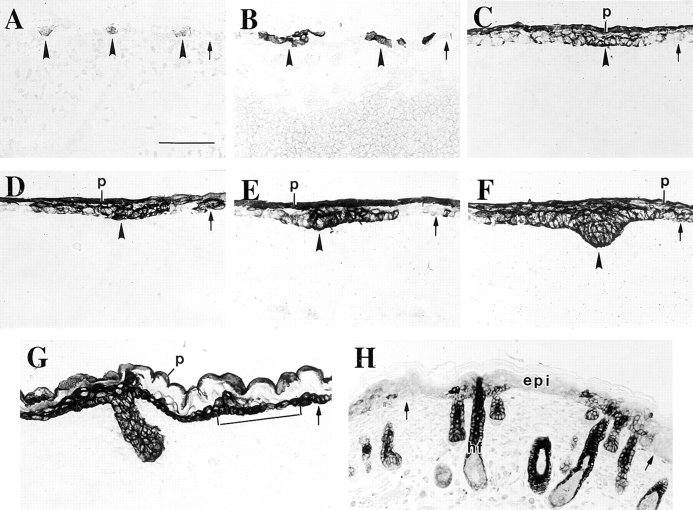
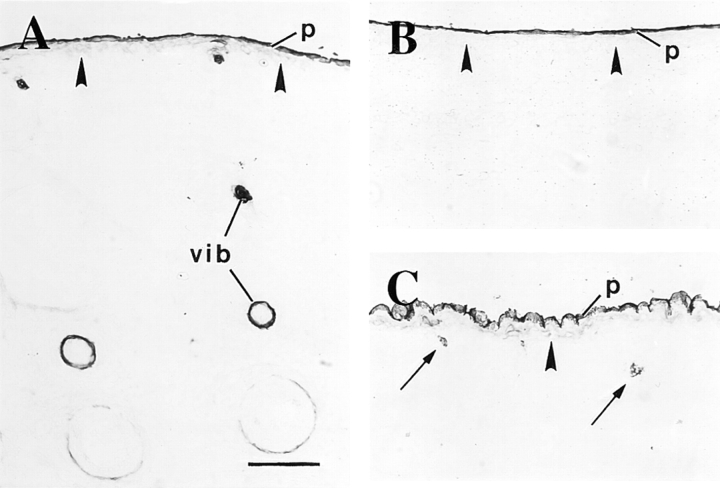
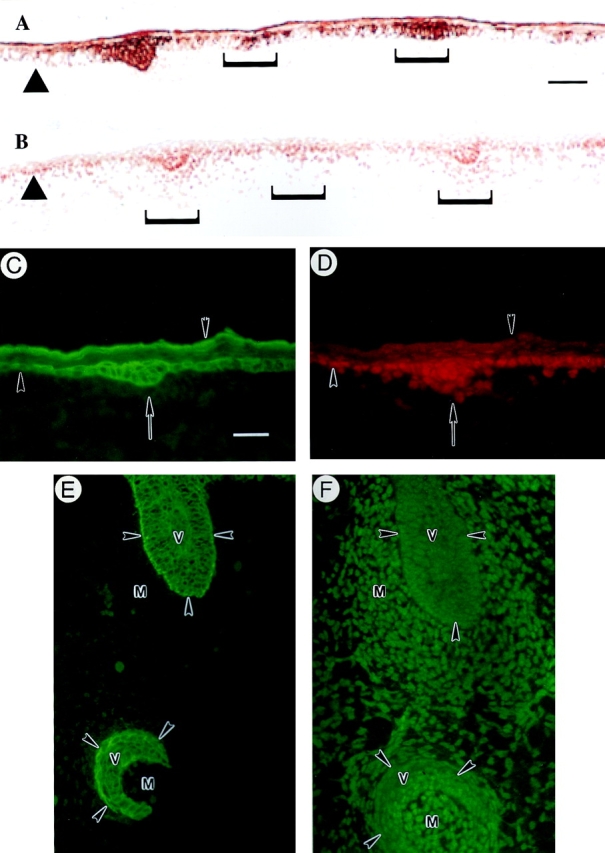
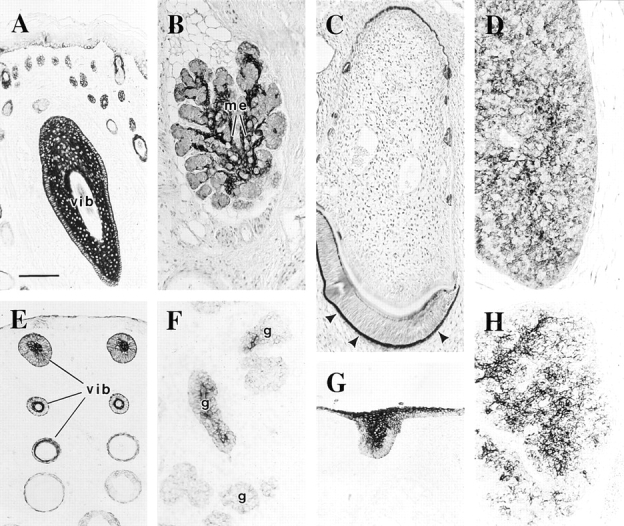
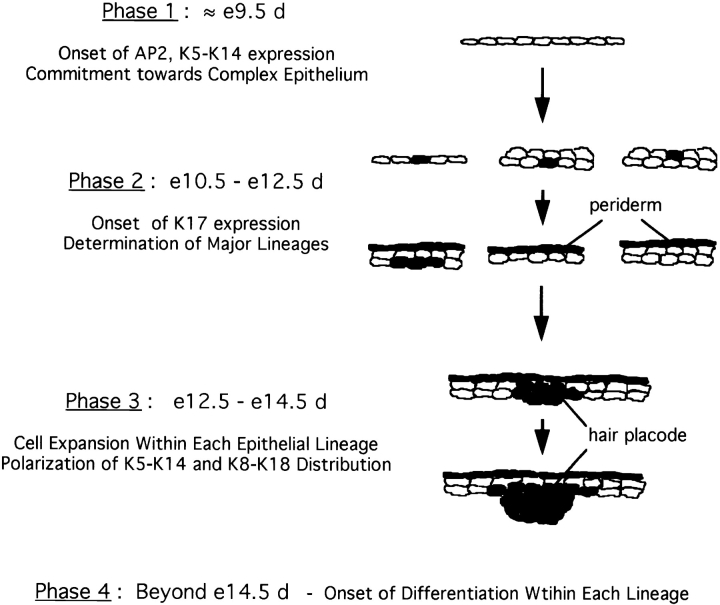
The type I keratin 17 (K17) shows a peculiar localization in human epithelial appendages including hair follicles, which undergo a growth cycle throughout adult life. Additionally K17 is induced, along with K6 and K16, early after acute injury to human skin. To gain further insights into its potential function(s), we cloned the mouse K17 gene and investigated its expression during skin development. Synthesis of K17 protein first occurs in a subset of epithelial cells within the single-layered, undifferentiated ectoderm of embryonic day 10.5 mouse fetuses. In the ensuing 48 h, K17-expressing cells give rise to placodes, the precursors of ectoderm-derived appendages (hair, glands, and tooth), and to periderm. During early development, there is a spatial correspondence in the distribution of K17 and that of lymphoid-enhancer factor (lef-1), a DNA-bending protein involved in inductive epithelial-mesenchymal interactions. We demonstrate that ectopic lef-1 expression induces K17 protein in the skin of adult transgenic mice. The pattern of K17 gene expression during development has direct implications for the morphogenesis of skin epithelia, and points to the existence of a molecular relationship between development and wound repair.
Figures

References
-
- Bessho Y, Nakanishi S, Nawa H. Glutamate receptor agonists enhance the expression of BDNF mRNA in cultured cerebellar granule cells. Mol Brain Res. 1993;18:201–208. - PubMed
-
- Bonnekoh B, Huerkamp C, Wevers A, Geisel J, Sebok B, Bange FC, Greenhalgh DA, Bottger EC, Krieg T, Mahrle G. Up-regulation of keratin 17 expression in human HaCaT keratinocytes by interferon-gamma. J Invest Dermatol. 1995;104:58–61. - PubMed
-
- Boulikas T. A compilation and classification of DNA binding sites for protein transcription factors from vertebrates. Crit Rev Eukaryot Gene Expr. 1994;4:117–322. - PubMed
-
- Bousquet O, Coulombe PA. Cytoskeleton: missing links found? . Curr Biol. 1996;6:1563–1566. - PubMed
-
- Brock J, McCluskey J, Baribault H, Martin P. Perfect wound healing in the keratin 8 deficient mouse embryo. Cell Motil Cytoskeleton. 1996;35:358–366. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
