

Contributions of pathway and neuron to preferential motor reinnervation
- PMID: 9786974
- PMCID: PMC6793544
- DOI: 10.1523/JNEUROSCI.18-21-08674.1998
Contributions of pathway and neuron to preferential motor reinnervation
Abstract
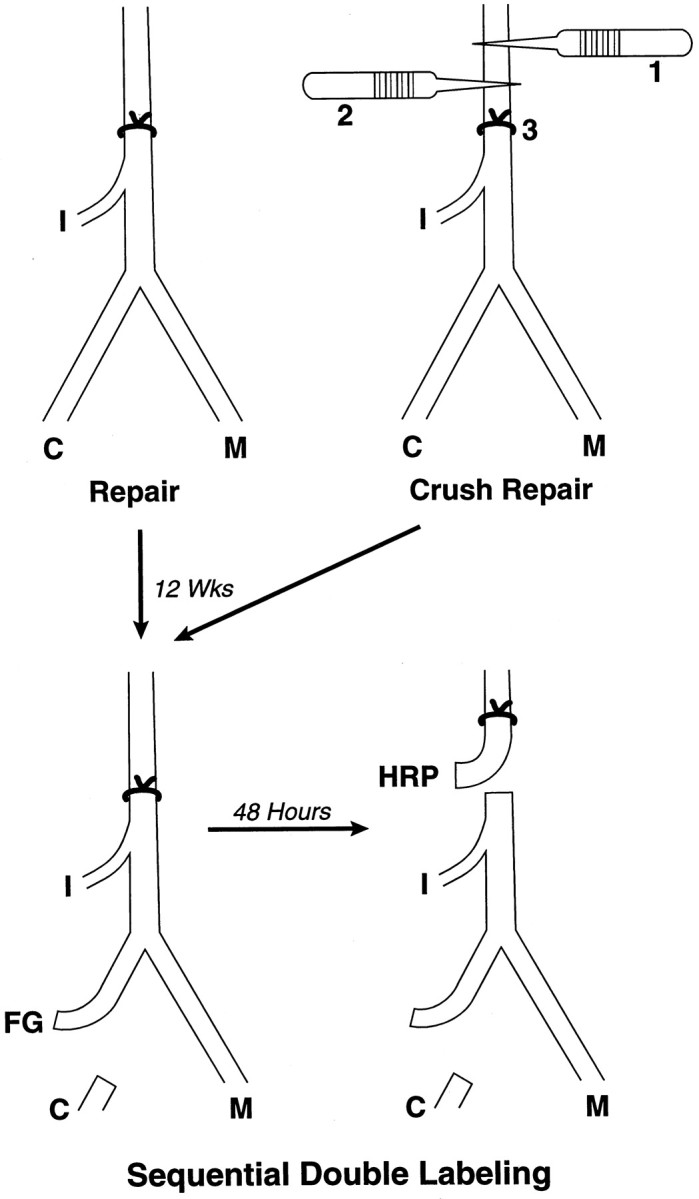
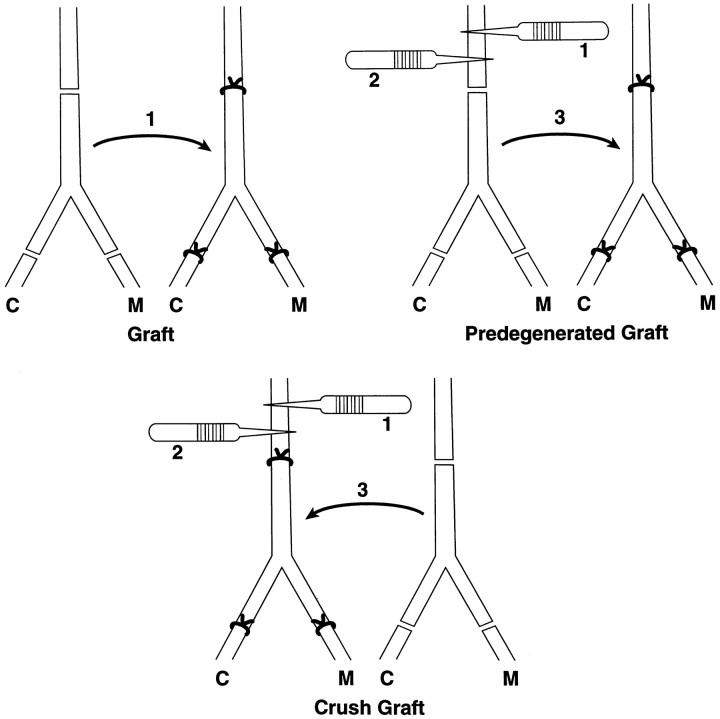
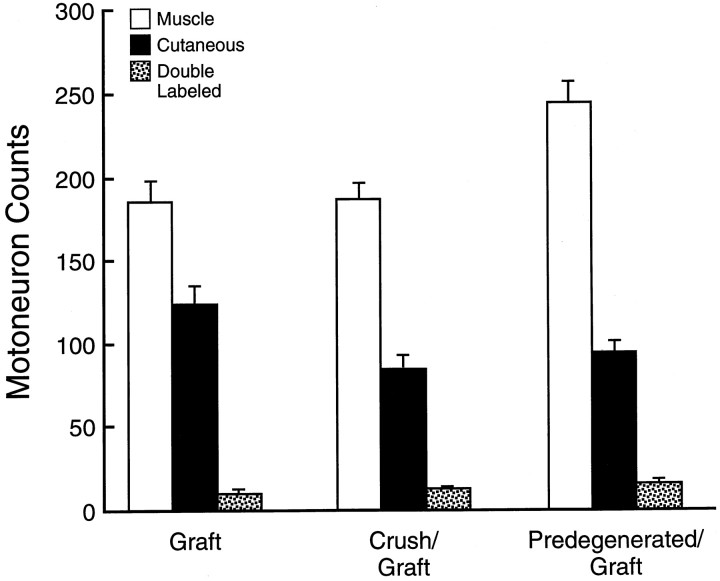
Motor axons regenerating after transection of mixed nerve preferentially reinnervate distal muscle branches, a process termed preferential motor reinnervation (PMR). Motor axon collaterals appear to enter both cutaneous and muscle Schwann cell tubes on a random basis. Double-labeling studies suggest that PMR is generated by pruning collaterals from cutaneous pathways while maintaining those in motor pathways (the "pruning hypothesis"). If all collaterals projecting to muscle are saved, then stimulation of regenerative sprouting should increase specificity by increasing the number of motoneurons with at least one collateral in a muscle pathway. In the current experiments, collateral sprouting is stimulated by crushing the nerve proximal to the repair site before suture, a maneuver that also conditions the neuron and predegenerates the distal pathway. Control experiments are performed to separate these effects from those of collateral generation. Experiments were performed on the rat femoral nerve and evaluated by exposing its terminal cutaneous and muscle branches to HRP or Fluoro-Gold. Crush proximal to the repair site increased motor axon collaterals at least fivefold and significantly increased the percentage of correctly projecting motoneurons, consistent with the pruning hypothesis. Conditioning the nerve with distal crushes before repair had no effect on specificity. A graft model was used to separate the effects of collateral generation and distal stump predegeneration. Previous crush of the proximal femoral nerve significantly increased the specificity of fresh graft reinnervation. Stimulation of regenerative collateral sprouting thus increased PMR, confirming the pruning hypothesis. However, this effect was overshadowed by the dramatic specificity with which predegenerated grafts were reinnervated by fresh uncrushed proximal axons. These unexpected effects of predegeneration on specificity could involve a variety of possible mechanisms and warrant further study because of their mechanistic and clinical implications.
Figures



References
-
- Abercrombie M. Estimation of nuclear population from microtome sections. Anat Rec. 1946;94:239–247. - PubMed
-
- Baptista CA, Macagno ER. Modulation of the pattern of axonal projections of a leech motor neuron by ablation or transplantation of its target. Neuron. 1988;1:949–962. - PubMed
-
- Bedi K, Winter J, Berry M, Cohen J. Adult rat dorsal root ganglion neurons extend neurites on predegenerated but not on normal peripheral nerves in vitro. Eur J Neurosci. 1992;4:193–200. - PubMed
-
- Bentley FH, Hill M. Nerve grafting. Br J Surg. 1936;24:368–387.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources