

A conventional myosin motor drives neurite outgrowth
- PMID: 9789024
- PMCID: PMC23673
- DOI: 10.1073/pnas.95.22.12967
A conventional myosin motor drives neurite outgrowth
Abstract
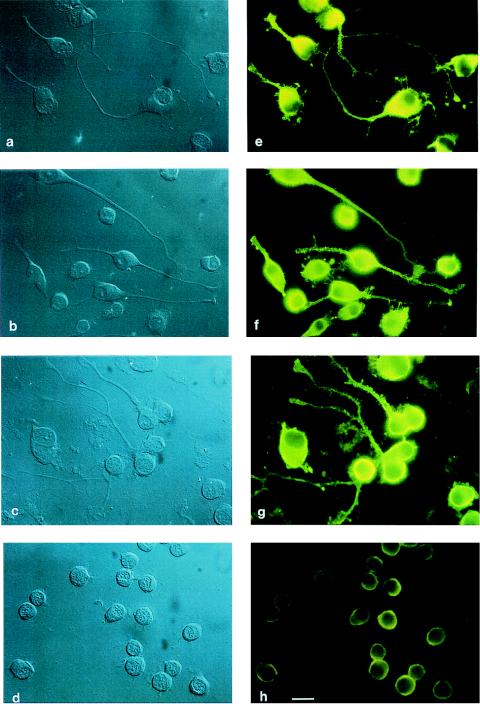
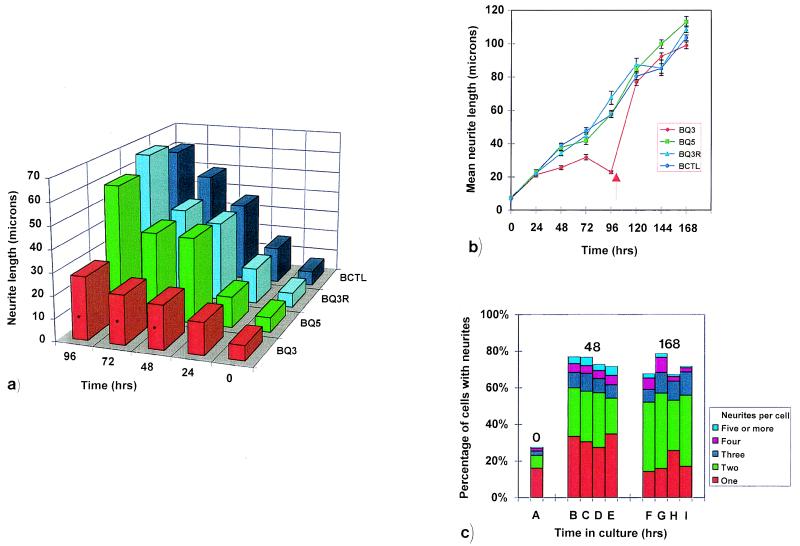
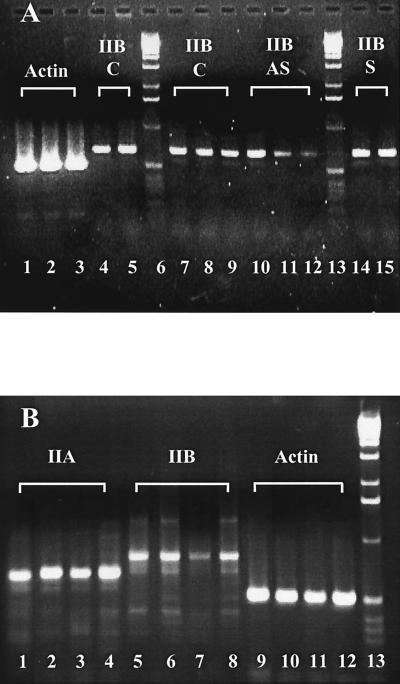
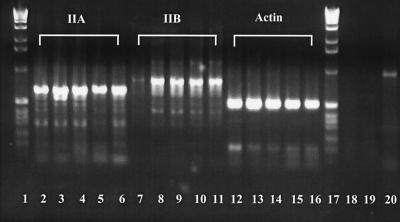
Neuritic outgrowth is a striking example of directed motility, powered through the actions of molecular motors. Members of the myosin superfamily of actin-associated motors have been implicated in this complex process. Although conventional myosin II is known to be present in neurons, where it is localized at the leading edge of growth cones and in the cell cortex close to the plasma membrane, its functional involvement in growth cone motility has remained unproven. Here, we show that antisense oligodeoxyribonucleotides, complementary to a specific isoform of conventional myosin (myosin IIB), attenuate filopodial extension whereas sense and scrambled control oligodeoxyribonucleotides have no effect. Attenuation is shown to be reversible, neurite outgrowth being restored after cessation of the antisense regimen. Myosin IIB mRNA was present during active neurite extension, but levels were minimal in phenotypically rounded cells before neurite outgrowth and message levels decreased during antisense treatment. By contrast, the myosin IIA isoform is shown to be expressed constitutively both before and during neurite outgrowth and throughout exposure to myosin IIB antisense oligodeoxyribonucleotides. These results provide direct evidence that a conventional two-headed myosin is required for growth cone motility and is responsible, at least in part, for driving neuritic process outgrowth.
Figures
References
-
- Letourneau P C, Kater S B, Macagno E R, editors. The Nerve Growth Cone. New York: Raven; 1991.
-
- Puszkin S, Berl S, Puszkin E, Clarke D. Science. 1968;161:170–171. - PubMed
-
- Puszkin S, Nicklas W J, Berl S. J Neurochem. 1972;19:1319–1333. - PubMed
-
- Lewis A K, Bridgman P C. Cell Motil Cytoskeleton. 1996;33:130–150. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
