

Ionizing radiation and short wavelength UV activate NF-kappaB through two distinct mechanisms
- PMID: 9789032
- PMCID: PMC23690
- DOI: 10.1073/pnas.95.22.13012
Ionizing radiation and short wavelength UV activate NF-kappaB through two distinct mechanisms
Abstract
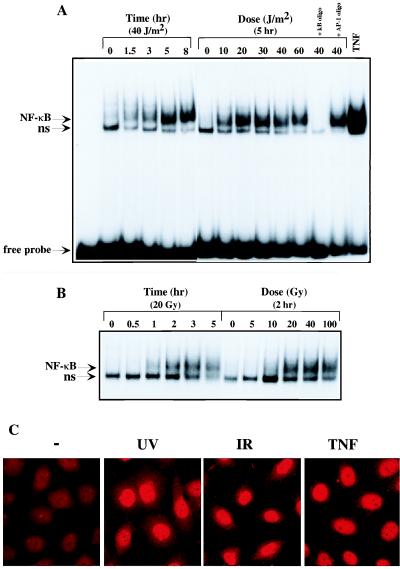
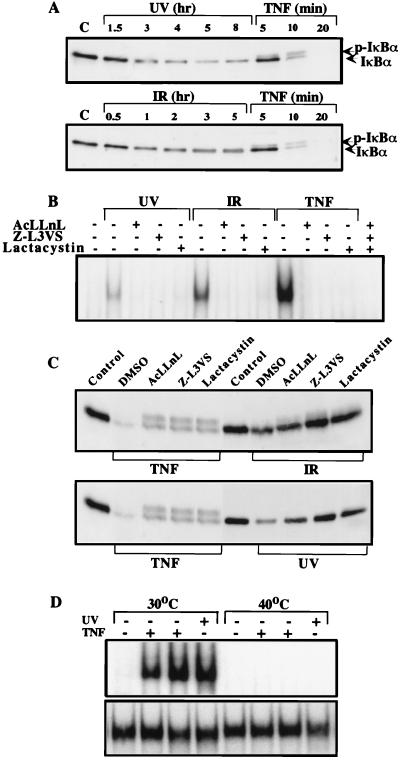
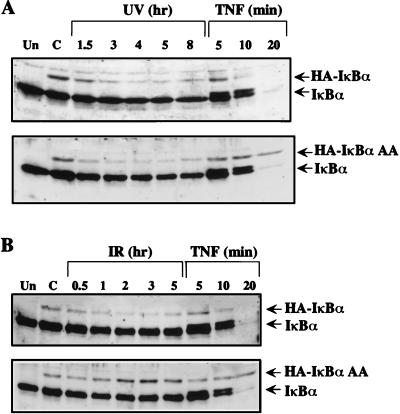
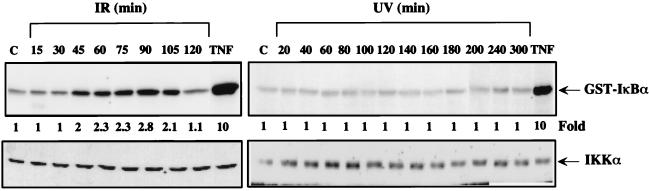
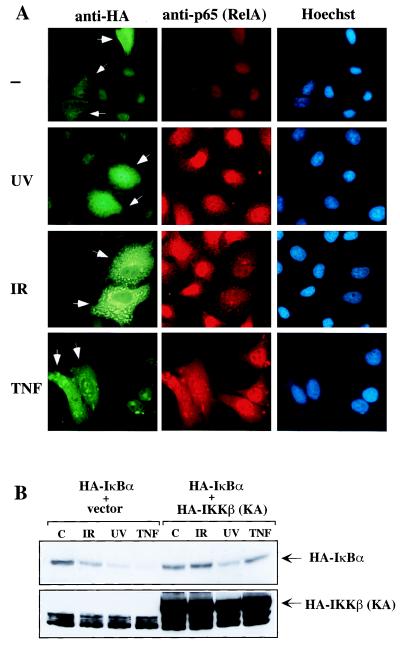
We examined the mechanisms by which two different types of photonic radiation, short wavelength UV (UV-C) and gamma radiation, activate transcription factor NF-kappaB. Exposure of mammalian cells to either form of radiation resulted in induction with similar kinetics of NF-kappaB DNA binding activity, nuclear translocation of its p65(RelA) subunit, and degradation of the major NF-kappaB inhibitor IkappaBalpha. In both cases, induction of NF-kappaB activity was attenuated by proteasome inhibitors and a mutation in ubiquitin-activating enzyme, suggesting that both UV-C and gamma radiation induce degradation of IkappaBs by means of the ubiquitin/proteasome pathway. However, although the induction of IkappaBalpha degradation by gamma rays was dependent on its phosphorylation at Ser-32 and Ser-36, UV-C-induced IkappaBalpha degradation was not dependent on phosphorylation of these residues. Even the "super repressor" IkappaBalpha mutant, which contains alanines at positions 32 and 36, was still susceptible to UV-C-induced degradation. Correspondingly, we found that gamma radiation led to activation of IKK, the protein kinase that phosphorylates IkappaBalpha at Ser-32 and Ser-36, whereas UV-C radiation did not. Furthermore, expression of a catalytically inactive IKKbeta mutant prevented NF-kappaB activation by gamma radiation, but not by UV-C. These results indicate that gamma radiation and UV-C activate NF-kappaB through two distinct mechanisms.
Figures
References
-
- Herrlich P, Ponta H, Rahmsdorf H J. Rev Physiol Biochem Pharmacol. 1992;119:187–223. - PubMed
-
- Canman C E, Kastan M B. Nature (London) 1996;384:213–214. - PubMed
-
- Ko L J, Prives C. Genes Dev. 1996;10:1054–1072. - PubMed
-
- Liu Z-G, Baskaran R, Lea-Chou E T, Wood L D, Chen Y, Karin M, Wang J Y J. Nature (London) 1996;384:273–276. - PubMed
-
- El-Deiry W S, Tokino T, Velculescu V E, Levy D B, Parsons R, Trent J M, Lin D, Mercer W E, Kinzler K W, Vogelstein B. Cell. 1993;75:817–825. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
