

Inhibition of arachidonate 5-lipoxygenase triggers massive apoptosis in human prostate cancer cells
- PMID: 9789062
- PMCID: PMC23752
- DOI: 10.1073/pnas.95.22.13182
Inhibition of arachidonate 5-lipoxygenase triggers massive apoptosis in human prostate cancer cells
Abstract
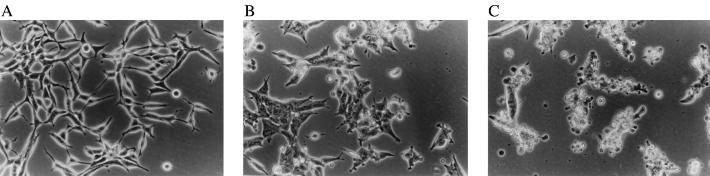
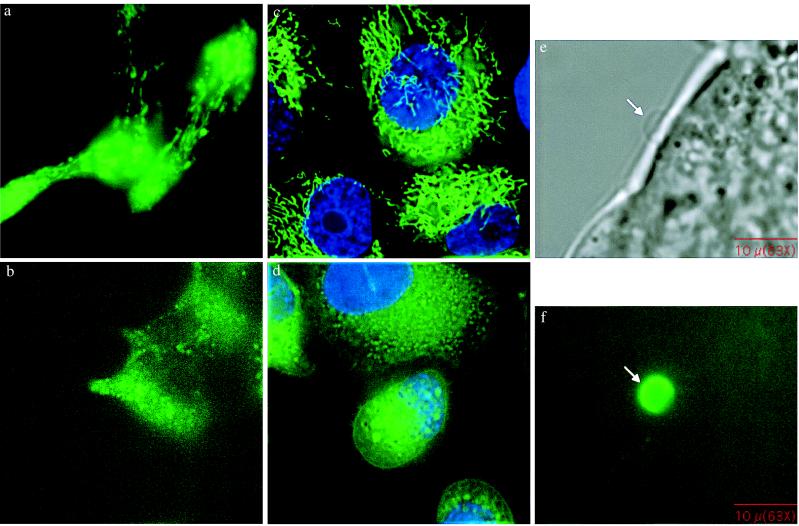
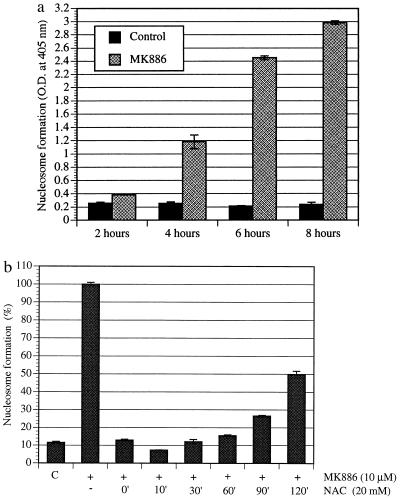
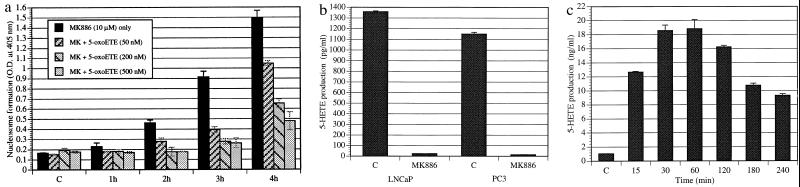
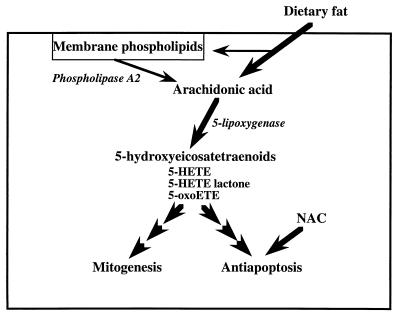
Diets high in fat are associated with an increased risk of prostate cancer, although the molecular mechanism is still unknown. We have previously reported that arachidonic acid, an omega-6 fatty acid common in the Western diet, stimulates proliferation of prostate cancer cells through production of the 5-lipoxygenase metabolite, 5-HETE (5-hydroxyeicosatetraenoic acid). We now show that 5-HETE is also a potent survival factor for human prostate cancer cells. These cells constitutively produce 5-HETE in serum-free medium with no added stimulus. Exogenous arachidonate markedly increases the production of 5-HETE. Inhibition of 5-lipoxygenase by MK886 completely blocks 5-HETE production and induces massive apoptosis in both hormone-responsive (LNCaP) and -nonresponsive (PC3) human prostate cancer cells. This cell death is very rapid: cells treated with MK886 showed mitochondrial permeability transition between 30 and 60 min, externalization of phosphatidylserine within 2 hr, and degradation of DNA to nucleosomal subunits beginning within 2-4 hr posttreatment. Cell death was effectively blocked by the thiol antioxidant, N-acetyl-L-cysteine, but not by androgen, a powerful survival factor for prostate cancer cells. Apoptosis was specific for 5-lipoxygenase-programmed cell death was not observed with inhibitors of 12-lipoxygenase, cyclooxygenase, or cytochrome P450 pathways of arachidonic acid metabolism. Exogenous 5-HETE protects these cells from apoptosis induced by 5-lipoxygenase inhibitors, confirming a critical role of 5-lipoxygenase activity in the survival of these cells. These findings provide a possible molecular mechanism by which dietary fat may influence the progression of prostate cancer.
Figures
References
-
- Parker S L, Tong T, Bolden S, Wingo P A. Ca Cancer J Clin. 1997;47:5–27. - PubMed
-
- Crawford E D, Eisenberger M A, McLeod D G, Spaulding J T, Benson R, Dorr A, Blumenstein B A, Davis M A, Goodman P J. N Engl J Med. 1989;321:419–424. - PubMed
-
- Gann P H, Hennekens C H, Sacks F M, Grodstein F, Giovannucci E L, Stampfer M J. J Natl Cancer Inst. 1994;86:281–286. - PubMed
-
- West D W, Slattery M L, Robison L M, French T K, Mahoney A W. Cancer Causes Control. 1991;2:85–94. - PubMed
-
- Giovannucci E, Rimm E B, Colditz G A, Stampfer M J, Ascherio A, Chute C C, Willett W C. J Natl Cancer Inst. 1993;85:1571–1579. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
