

Ca2+/calmodulin-dependent kinase II mediates simultaneous enhancement of gap-junctional conductance and glutamatergic transmission
- PMID: 9789078
- PMCID: PMC23780
- DOI: 10.1073/pnas.95.22.13272
Ca2+/calmodulin-dependent kinase II mediates simultaneous enhancement of gap-junctional conductance and glutamatergic transmission
Abstract
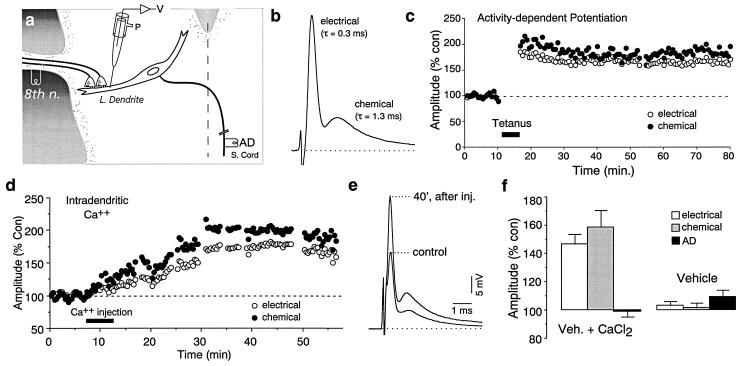
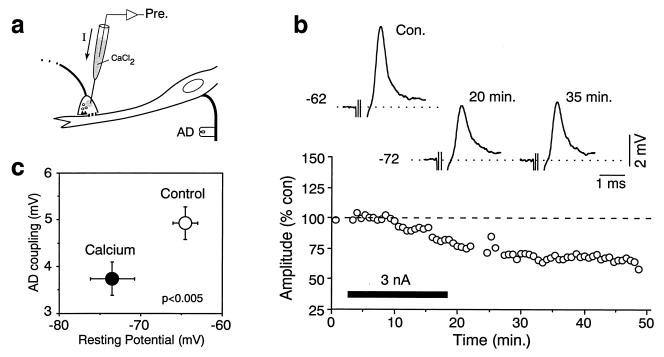
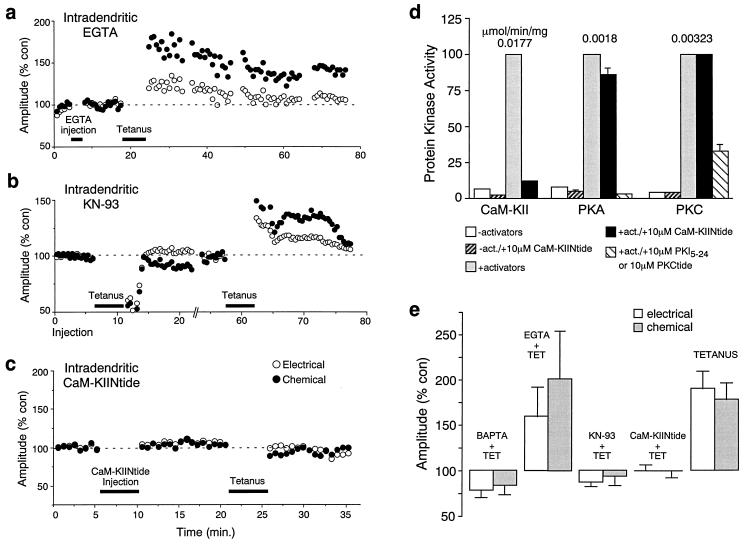
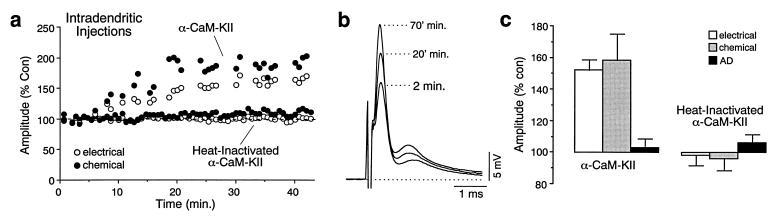
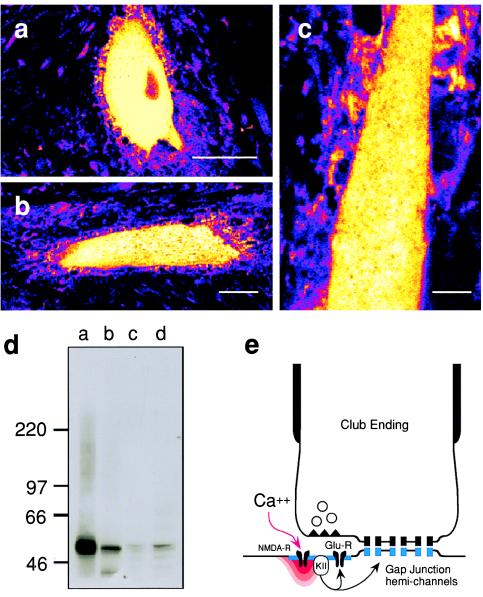
While chemical synapses are very plastic and modifiable by defined activity patterns, gap junctions, which mediate electrical transmission, have been classically perceived as passive intercellular channels. Excitatory transmission between auditory afferents and the goldfish Mauthner cell is mediated by coexisting gap junctions and glutamatergic synapses. Although an increased intracellular Ca2+ concentration is expected to reduce gap junctional conductance, both components of the synaptic response were instead enhanced by postsynaptic increases in Ca2+ concentration, produced by patterned synaptic activity or intradendritic Ca2+ injections. The synaptically induced potentiations were blocked by intradendritic injection of KN-93, a Ca2+/calmodulin-dependent kinase (CaM-K) inhibitor, or CaM-KIINtide, a potent and specific peptide inhibitor of CaM-KII, whereas the responses were potentiated by injection of an activated form of CaM-KII. The striking similarities of the mechanisms reported here with those proposed for long-term potentiation of mammalian glutamatergic synapses suggest that gap junctions are also similarly regulated and indicate a primary role for CaM-KII in shaping and regulating interneuronal communication, regardless of its modality.
Figures
References
-
- Malenka R C, Nicoll R A. Trends Neurosci. 1993;16:521–527. - PubMed
-
- Bennett M V L. In: Cellular Biology of Neurons, Handbook of Physiology. Kandel E R, editor. 1, Sect. I. Baltimore: Williams and Wilkins; 1977. pp. 357–416.
-
- Yang X D, Korn H, Faber D S. Nature (London) 1990;348:542–545. - PubMed
-
- Bartelmez G W. J Comp Neurol. 1915;25:87–128.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
