

A G protein gamma subunit-like domain shared between RGS11 and other RGS proteins specifies binding to Gbeta5 subunits
- PMID: 9789084
- PMCID: PMC23793
- DOI: 10.1073/pnas.95.22.13307
A G protein gamma subunit-like domain shared between RGS11 and other RGS proteins specifies binding to Gbeta5 subunits
Abstract
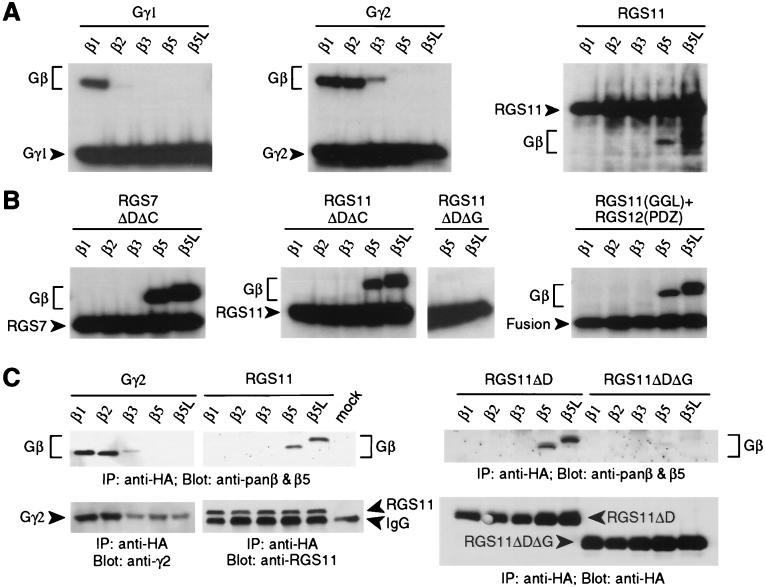
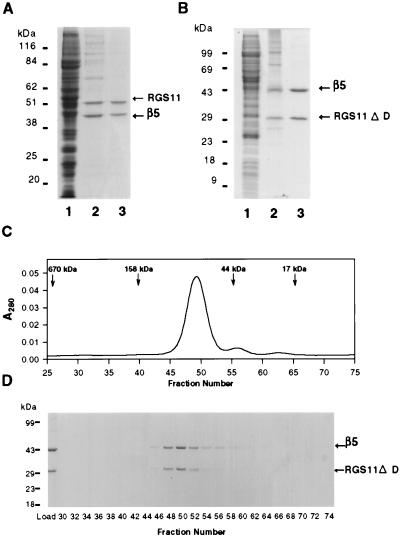
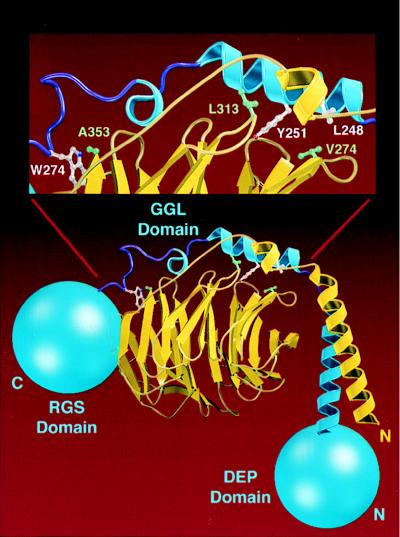
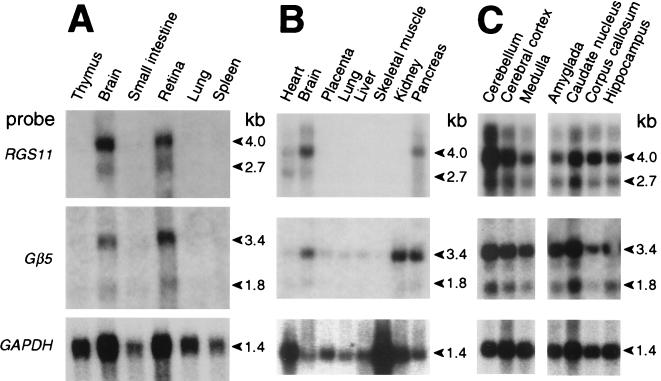
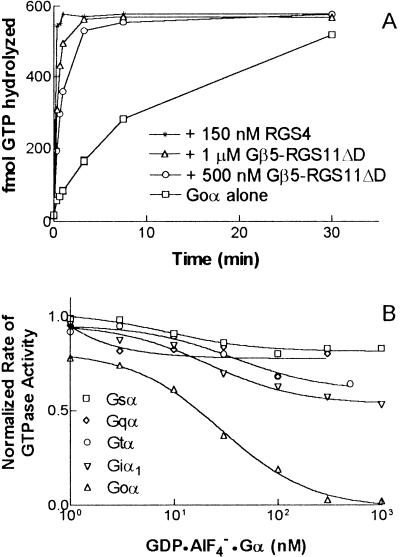
Regulators of G protein signaling (RGS) proteins act as GTPase-activating proteins (GAPs) toward the alpha subunits of heterotrimeric, signal-transducing G proteins. RGS11 contains a G protein gamma subunit-like (GGL) domain between its Dishevelled/Egl-10/Pleckstrin and RGS domains. GGL domains are also found in RGS6, RGS7, RGS9, and the Caenorhabditis elegans protein EGL-10. Coexpression of RGS11 with different Gbeta subunits reveals specific interaction between RGS11 and Gbeta5. The expression of mRNA for RGS11 and Gbeta5 in human tissues overlaps. The Gbeta5/RGS11 heterodimer acts as a GAP on Galphao, apparently selectively. RGS proteins that contain GGL domains appear to act as GAPs for Galpha proteins and form complexes with specific Gbeta subunits, adding to the combinatorial complexity of G protein-mediated signaling pathways.
Figures

References
-
- Dohlman H G, Thorner J. J Biol Chem. 1997;272:3871–3874. - PubMed
-
- Koelle M R. Curr Opin Cell Biol. 1997;9:143–147. - PubMed
-
- Berman D M, Gilman A G. J Biol Chem. 1998;273:1269–1272. - PubMed
-
- Snow B E, Hall R A, Krumins A M, Brothers G M, Bouchard D, Brothers C A, Chung S, Mangion J, Gilman A G, Lefkowitz R J, Siderovski D P. J Biol Chem. 1998;273:17749–17755. - PubMed
-
- Kozasa T, Jiang X, Hart M J, Sternweis P M, Singer W D, Gilman A G, Bollag G, Sternweis P C. Science. 1998;280:2109–2111. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
