

Identification of chlorobenzene dioxygenase sequence elements involved in dechlorination of 1,2,4,5-tetrachlorobenzene
- PMID: 9791099
- PMCID: PMC107608
- DOI: 10.1128/JB.180.21.5520-5528.1998
Identification of chlorobenzene dioxygenase sequence elements involved in dechlorination of 1,2,4,5-tetrachlorobenzene
Abstract
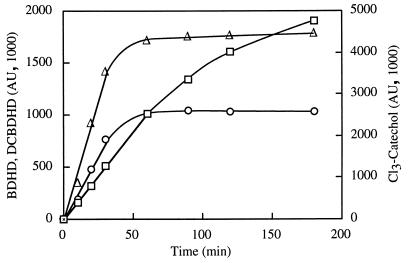
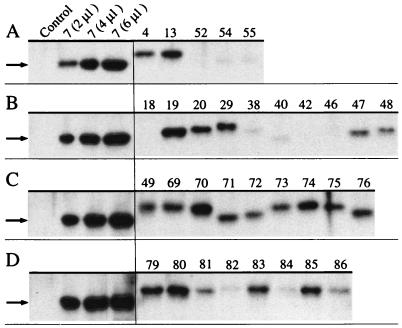
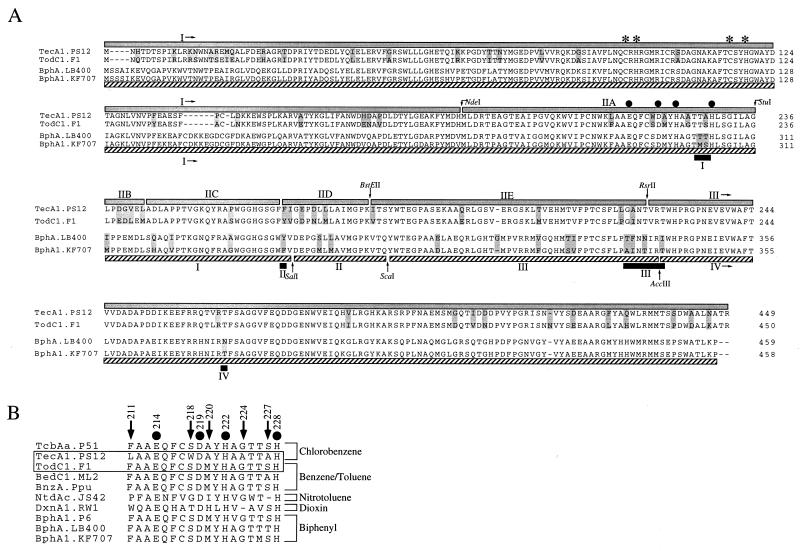
The TecA chlorobenzene dioxygenase and the TodCBA toluene dioxygenase exhibit substantial sequence similarity yet have different substrate specificities. Escherichia coli cells producing recombinant TecA enzyme dioxygenate and simultaneously eliminate a halogen substituent from 1,2,4,5-tetrachlorobenzene but show no activity toward benzene, whereas those producing TodCBA dioxygenate benzene but not tetrachlorobenzene. A hybrid TecA dioxygenase variant containing the large alpha-subunit of the TodCBA dioxygenase exhibited a TodCBA dioxygenase specificity. Acquisition of dehalogenase activity was achieved by replacement of specific todC1 alpha-subunit subsequences by equivalent sequences of the tecA1 alpha-subunit. Substrate transformation specificities and rates by E. coli resting cells expressing hybrid systems were analyzed by high-performance liquid chromatography. This allowed the identification of both a single amino acid and potentially interacting regions required for dechlorination of tetrachlorobenzene. Hybrids with extended substrate ranges were generated that exhibited activity toward both benzene and tetrachlorobenzene. The regions determining substrate specificity in (chloro)benzene dioxygenases appear to be different from those previously identified in biphenyl dioxygenases.
Figures

References
-
- Asturias J A, Diaz E, Timmis K N. The evolutionary relationship of biphenyl dioxygenase from gram-positive Rhodococcus globerulus P6 to multicomponent dioxygenases from gram-negative bacteria. Gene. 1995;156:11–18. - PubMed
-
- Batie C J, Ballou D P, Correll C J. Phthalate dioxygenase reductase and related flavin-iron-sulphur containing electron transferases. In: Müller F, editor. Chemistry and biochemistry of flavoenzymes. Boca Raton, Fla: CRC Press; 1992. pp. 544–554.
-
- Beil S, Happe B, Timmis K N, Pieper D H. Genetic and biochemical characterization of the broad spectrum chlorobenzene dioxygenase from Burkholderia sp. strain PS12: dechlorination of 1,2,4,5-tetrachlorobenzene. Eur J Biochem. 1997;247:190–199. - PubMed
-
- Bradford M M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem. 1976;72:248–254. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
