

Mechanism of osmotic activation of the quaternary ammonium compound transporter (QacT) of Lactobacillus plantarum
- PMID: 9791101
- PMCID: PMC107610
- DOI: 10.1128/JB.180.21.5540-5546.1998
Mechanism of osmotic activation of the quaternary ammonium compound transporter (QacT) of Lactobacillus plantarum
Abstract
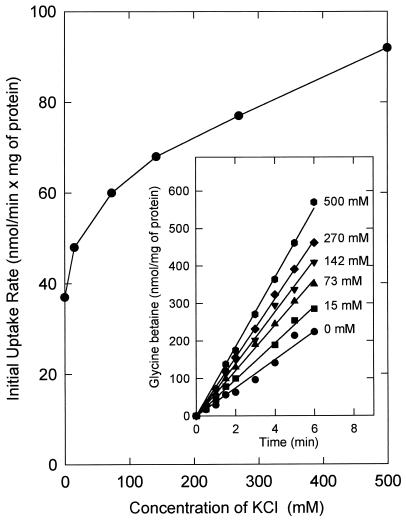
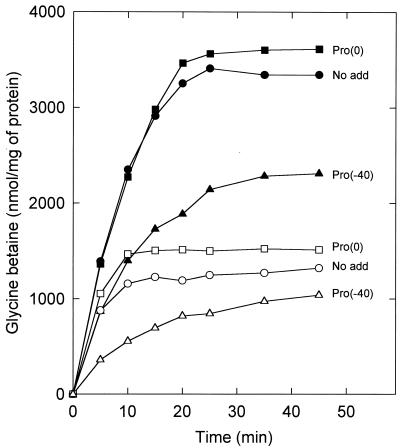
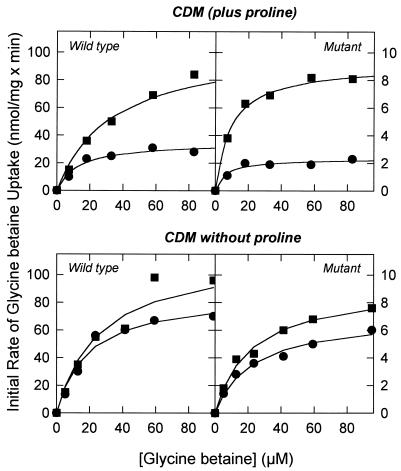
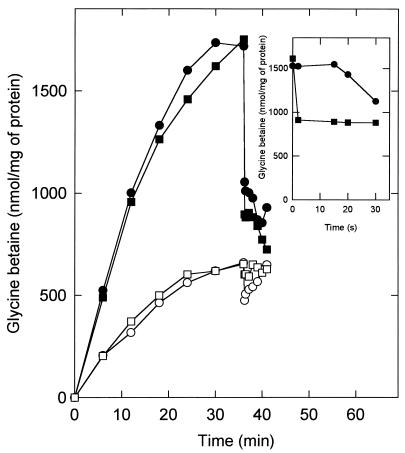
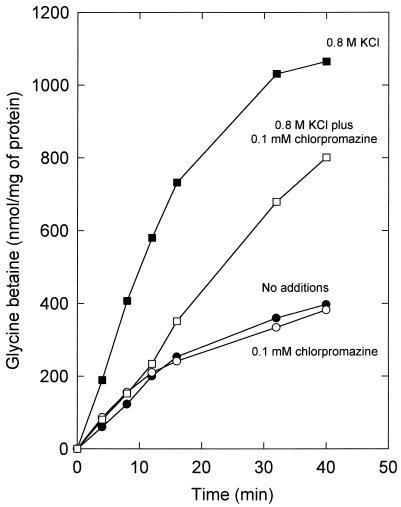
The accumulation of quaternary ammonium compounds in Lactobacillus plantarum is mediated via a single transport system with a high affinity for glycine betaine (apparent Km of 18 microM) and carnitine and a low affinity for proline (apparent Km of 950 microM) and other analogues. Mutants defective in the uptake of glycine betaine were generated by UV irradiation and selected on the basis of resistance to dehydroproline (DHP), a toxic proline analogue. Three independent DHP-resistant mutants showed reduced glycine betaine uptake rates and accumulation levels but behaved similarly to the wild type in terms of direct activation of uptake by high-osmolality conditions. Kinetic analysis of glycine betaine uptake and efflux in the wild-type and mutant cells is consistent with one uptake system for quaternary ammonium compounds in L. plantarum and a separate system(s) for their excretion. The mechanism of osmotic activation of the quaternary ammonium compound transport system (QacT) was studied. It was observed that the uptake rates were inhibited by the presence of internal substrate. Upon raising of the medium osmolality, the QacT system was rapidly activated (increase in maximal velocity) through a diminished inhibition by trans substrate as well as an effect that is independent of intracellular substrate. We also studied the effects of the cationic amphipath chlorpromazine, which inserts into the cytoplasmic membrane and thereby influences the uptake and efflux of glycine betaine. The results provide further evidence for the notion that the rapid efflux of glycine betaine upon osmotic downshock is mediated by a channel protein that is responding to membrane stretch or tension. The activation of QacT upon osmotic upshock seems to be brought about by a turgor-related parameter other than membrane stretch or tension.
Figures
References
-
- Faatz E, Middendorf A, Bremer E. Cloned structural genes for the osmotically regulated binding-protein-dependent glycine betaine transport system (ProU) of Escherichia coli K-12. Mol Microbiol. 1988;2:265–279. - PubMed
-
- Glaasker E, Konings W N, Poolman B. Glycine-betaine fluxes in Lactobacillus plantarum during osmostasis and hyper- and hypoosmotic shock. J Biol Chem. 1996;271:10060–10065. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
