

Molecular genetic analysis of phosphite and hypophosphite oxidation by Pseudomonas stutzeri WM88
- PMID: 9791102
- PMCID: PMC107611
- DOI: 10.1128/JB.180.21.5547-5558.1998
Molecular genetic analysis of phosphite and hypophosphite oxidation by Pseudomonas stutzeri WM88
Abstract
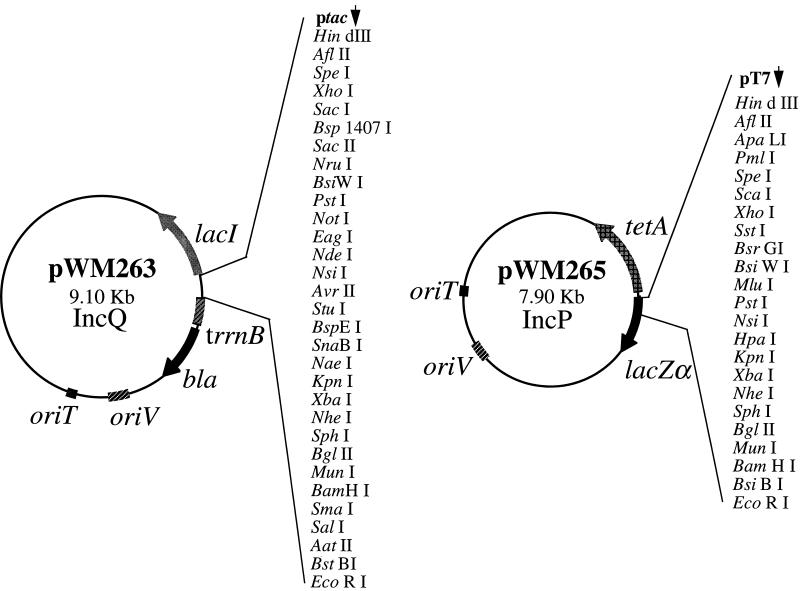
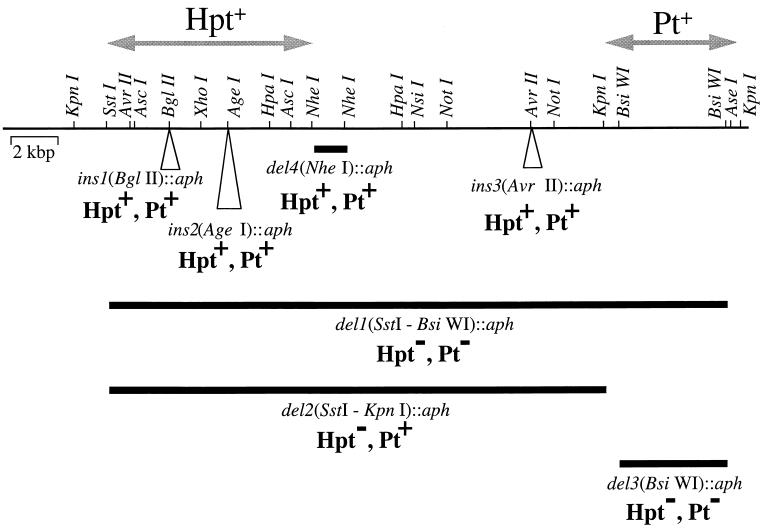
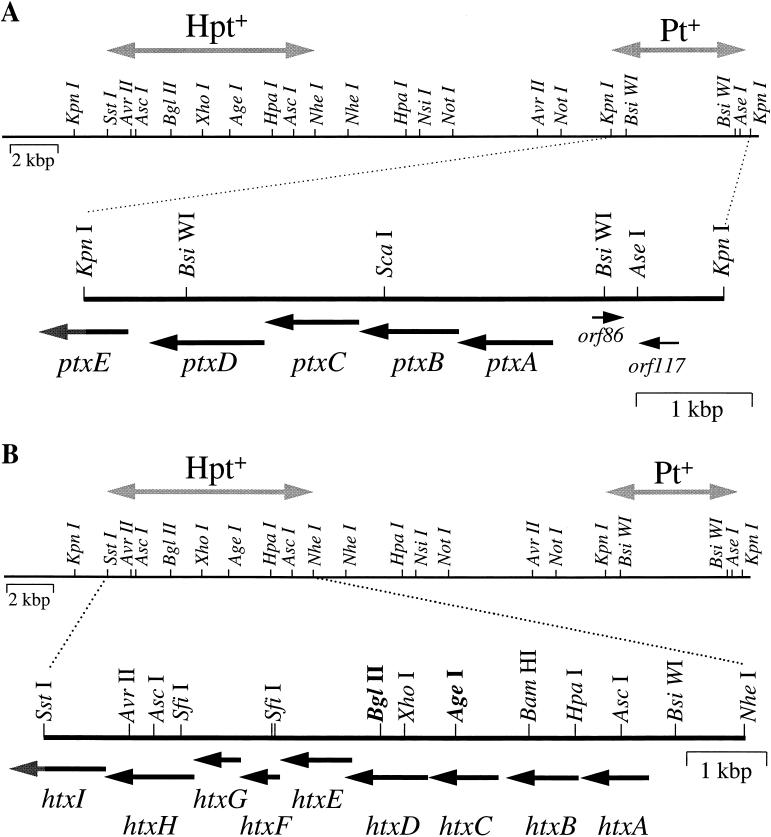
The first molecular and genetic characterization of a biochemical pathway for oxidation of the reduced phosphorus (P) compounds phosphite and hypophosphite is reported. The pathway was identified in Pseudomonas stutzeri WM88, which was chosen for detailed studies from a group of organisms isolated based on their ability to oxidize hypophosphite (+1 valence) and phosphite (+3 valence) to phosphate (+5 valence). The genes required for oxidation of both compounds by P. stutzeri WM88 were cloned on a single ca. 30-kbp DNA fragment by screening for expression in Escherichia coli and Pseudomonas aeruginosa. Two lines of evidence suggest that hypophosphite is oxidized to phosphate via a phosphite intermediate. First, plasmid subclones that conferred oxidation of phosphite, but not hypophosphite, upon heterologous hosts were readily obtained. All plasmid subclones that failed to confer phosphite oxidation also failed to confer hypophosphite oxidation. No subclones that conferred only hypophosphite expression were obtained. Second, various deletion derivatives of the cloned genes were made in vitro and recombined onto the chromosome of P. stutzeri WM88. Two phenotypes were displayed by individual mutants. Mutants with the region encoding phosphite oxidation deleted (based upon the subcloning results) lost the ability to oxidize either phosphite or hypophosphite. Mutants with the region encoding hypophosphite oxidation deleted lost only the ability to oxidize hypophosphite. The phenotypes displayed by these mutants also demonstrate that the cloned genes are responsible for the P oxidation phenotypes displayed by the original P. stutzeri WM88 isolate. The DNA sequences of the minimal regions implicated in oxidation of each compound were determined. The region required for oxidation of phosphite to phosphate putatively encodes a binding-protein-dependent phosphite transporter, an NAD+-dependent phosphite dehydrogenase, and a transcriptional activator of the lysR family. The region required for oxidation of hypophosphite to phosphite putatively encodes a binding-protein-dependent hypophosphite transporter and an alpha-ketoglutarate-dependent hypophosphite dioxygenase. The finding of genes dedicated to oxidation of reduced P compounds provides further evidence that a redox cycle for P may be important in the metabolism of this essential, and often growth-limiting, nutrient.
Figures

References
-
- Adams F, Conrad J P. Transition of phosphite to phosphate in soils. Soil Sci. 1953;75:361–371.
-
- Ausubel F M, Brent R, Kingston R E, Moore D D, Seidman J G, Smith J A, Struhl K. Current protocols in molecular biology. 1 and 2. New York, N.Y: John Wiley & Sons; 1992.
-
- Bennasar A, Rossello-Mora R, Lalucat J, Moore E R. 16S rRNA gene sequence analysis relative to genomovars of Pseudomonas stutzeri and proposal of Pseudomonas balearica sp. nov. Int J Syst Bacteriol. 1996;46:200–205. - PubMed
-
- Brosius J. Superpolylinkers in cloning and expression vectors. DNA. 1989;8:759–777. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
