

Gigantism in a bacterium, Epulopiscium fishelsoni, correlates with complex patterns in arrangement, quantity, and segregation of DNA
- PMID: 9791108
- PMCID: PMC107617
- DOI: 10.1128/JB.180.21.5601-5611.1998
Gigantism in a bacterium, Epulopiscium fishelsoni, correlates with complex patterns in arrangement, quantity, and segregation of DNA
Abstract
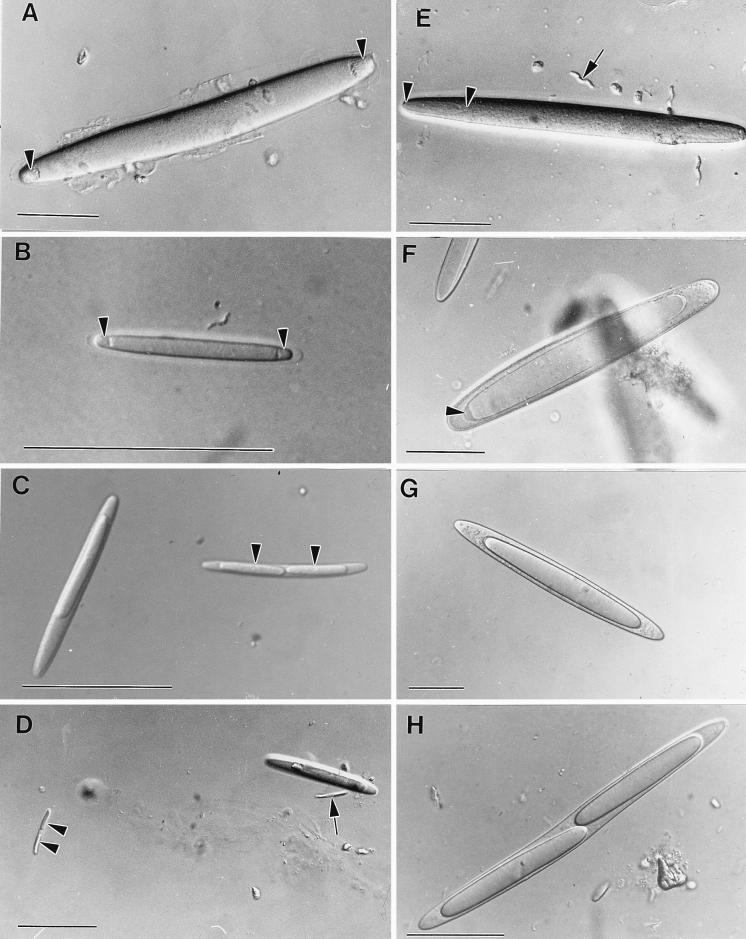
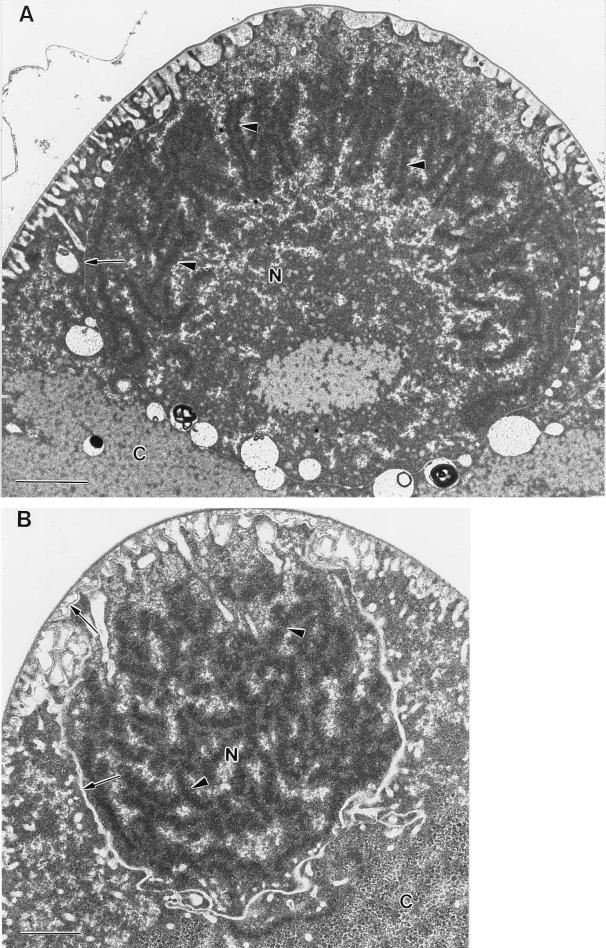
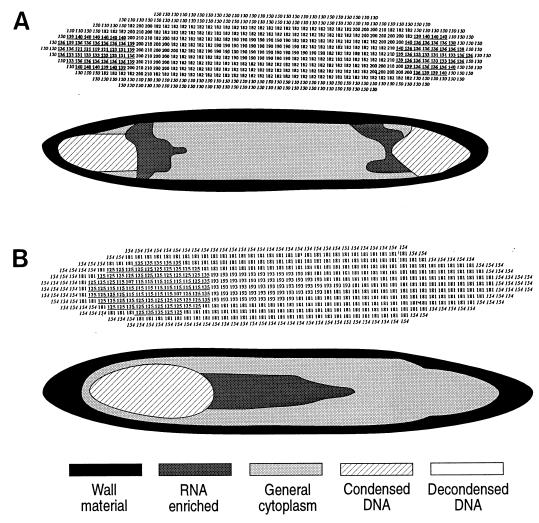
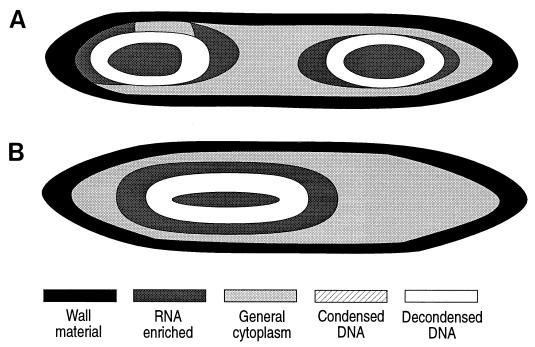
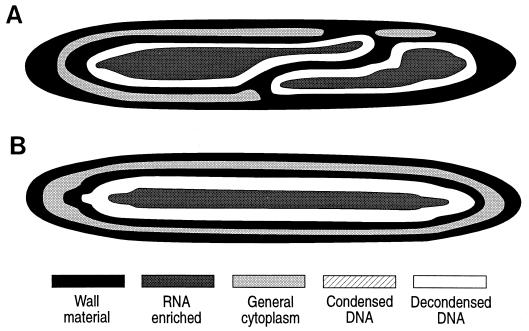
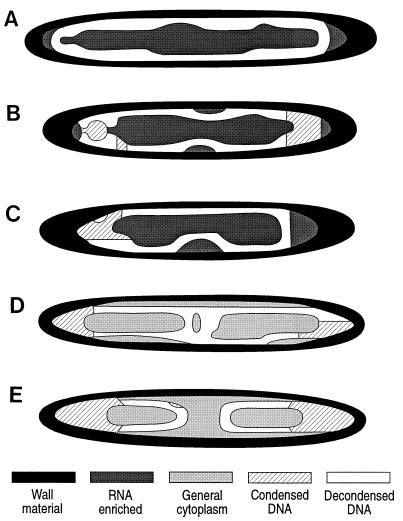
Epulopiscium fishelsoni, gut symbiont of the brown surgeonfish (Acanthurus nigrofuscus) in the Red Sea, attains a larger size than any other eubacterium, varies 10- to 20-fold in length (and >2, 000-fold in volume), and undergoes a complex daily life cycle. In early morning, nucleoids contain highly condensed DNA in elongate, chromosome-like structures which are physically separated from the general cytoplasm. Cell division involves production of two (rarely three) nucleoids within a cell, deposition of cell walls around expanded nucleoids, and emergence of daughter cells from the parent cell. Fluorescence measurements of DNA, RNA, and other cell components indicate the following. DNA quantity is proportional to cell volume over cell lengths of approximately 30 micrometers to >500 micrometers. For cells of a given size, nucleoids of cells with two nucleoids (binucleoid) contain approximately equal amounts of DNA. And each nucleoid of a binucleoid cell contains one-half the DNA of the single nucleoid in a uninucleoid cell of the same size. The life cycle involves approximately equal subdivision of DNA among daughter cells, formation of apical caps of condensed DNA from previously decondensed and diffusely distributed DNA, and "pinching" of DNA near the middle of the cell in the absence of new wall formation. Mechanisms underlying these patterns remain unclear, but formation of daughter nucleoids and cells occurs both during diurnal periods of host feeding and bacterial cell growth and during nocturnal periods of host inactivity when mean bacterial cell size declines.
Figures
References
-
- Angert E R, Clements K D, Pace N R. The largest bacterium. Nature. 1993;362:239–241. - PubMed
-
- Arndt-Jobin D J, Jobin T M. Fluorescence labeling and microscopy of DNA. Methods Cell Biol. 1989;30:417–448. - PubMed
-
- Bohm N, Sprenger E, Sandritter W. Absorbance and fluorescence cytometry of nuclear Fuelgen DNA. In: Thaer A A, Sernetz M, editors. Fluorescence techniques in cell biology. Berlin, Germany: Springer-Verlag; 1973.
-
- Bresler V, Fishelson L. Microfluorometrical study of benzo(a)pyrene and marker xenobiotics bioaccumulation in the bivalve Donax trunculus from clean and polluted sites along the Mediterranean shore of Israel. Dis Aquat Org. 1994;19:193–202.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
