

Neurotrimin mediates bifunctional effects on neurite outgrowth via homophilic and heterophilic interactions
- PMID: 9801370
- PMCID: PMC6792904
- DOI: 10.1523/JNEUROSCI.18-22-09312.1998
Neurotrimin mediates bifunctional effects on neurite outgrowth via homophilic and heterophilic interactions
Abstract
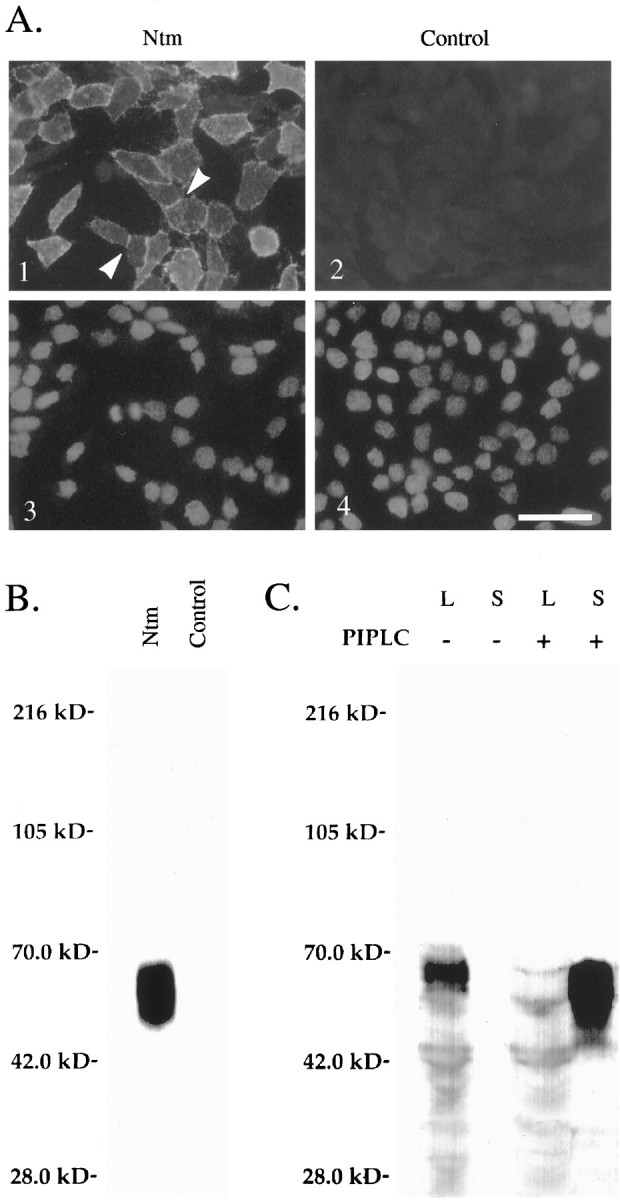
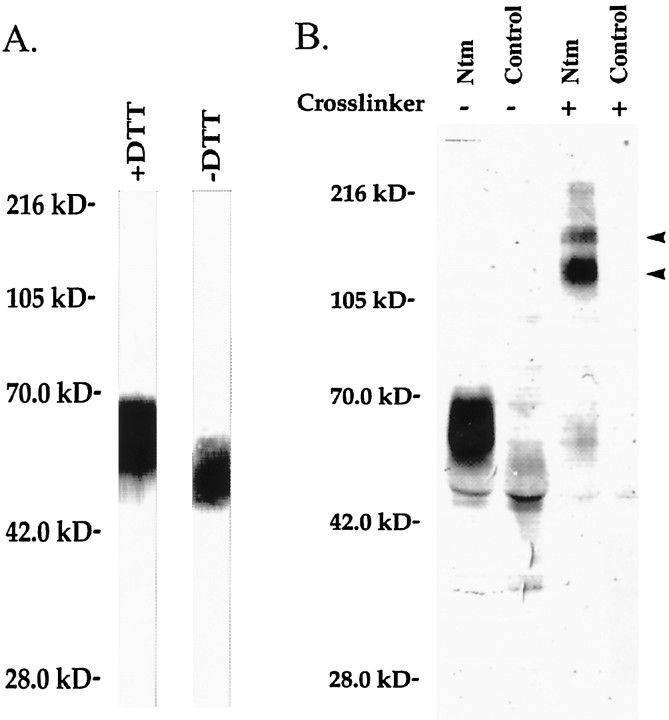
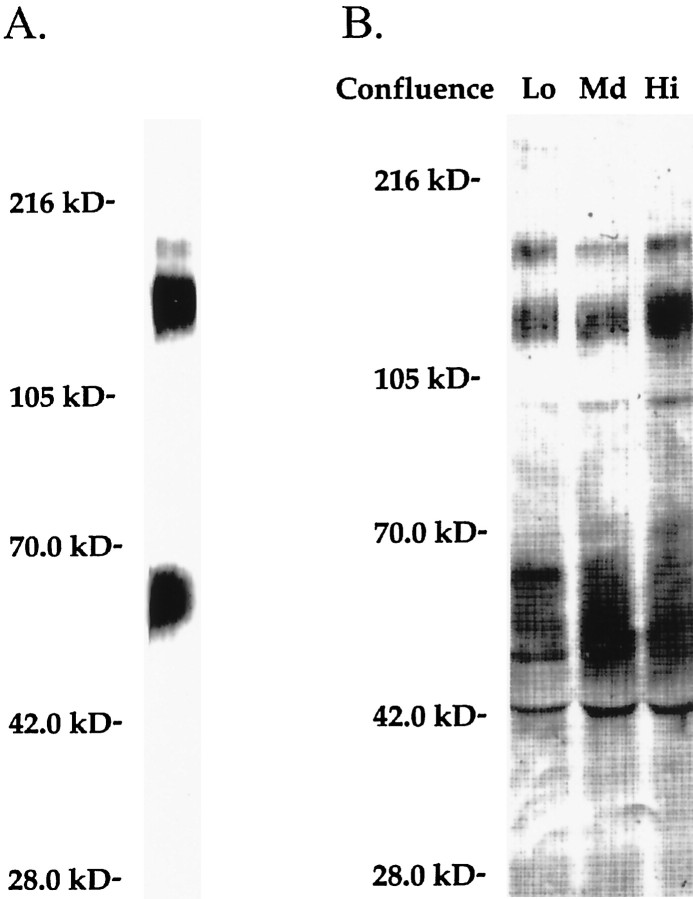
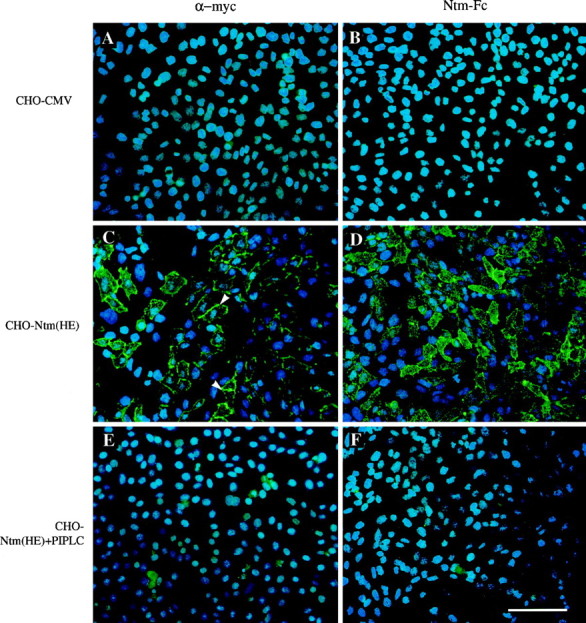
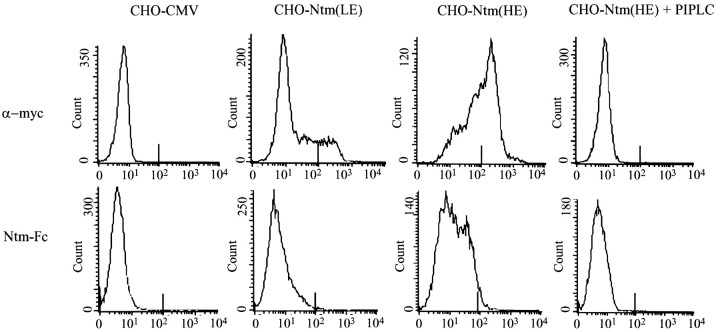
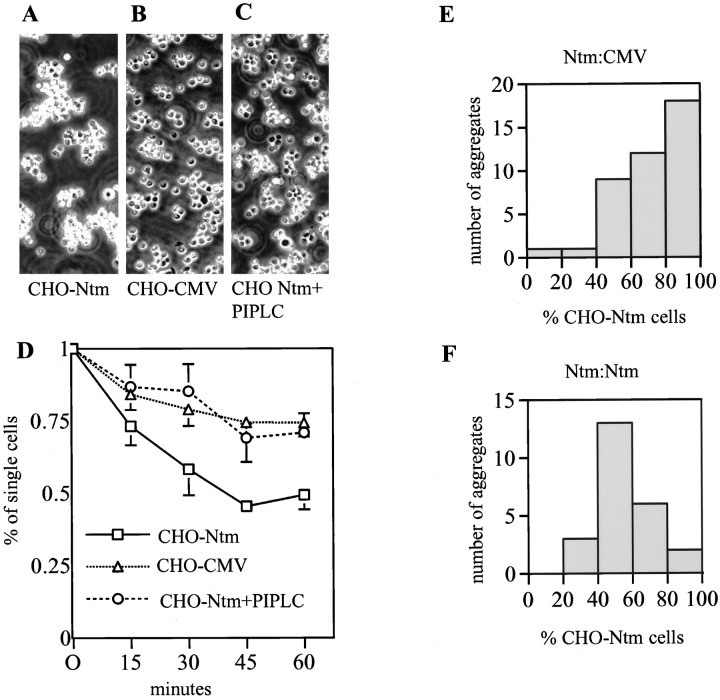
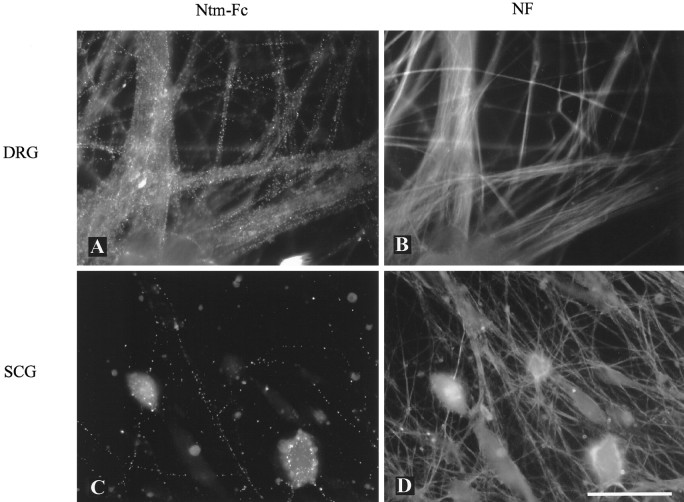
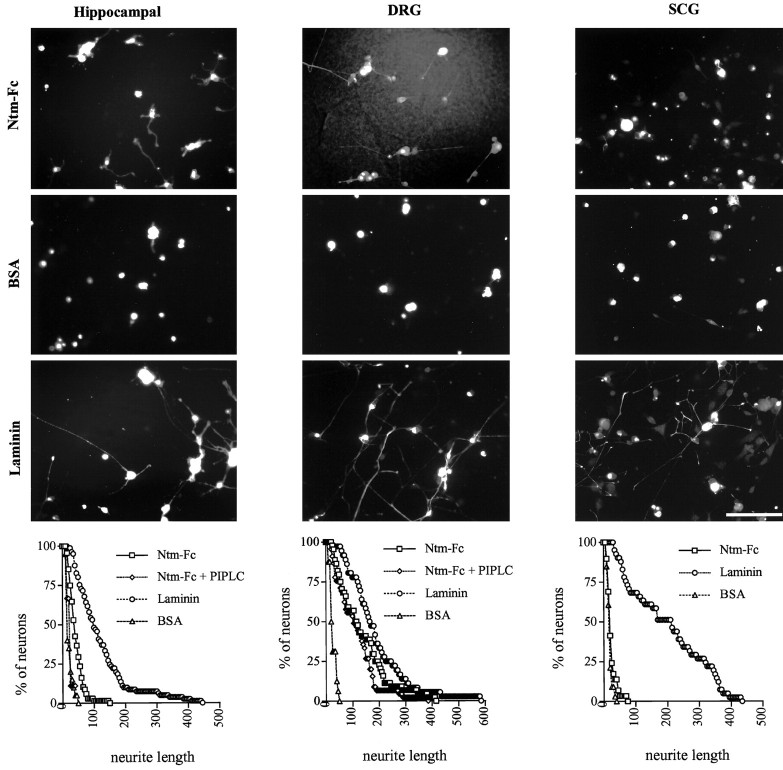
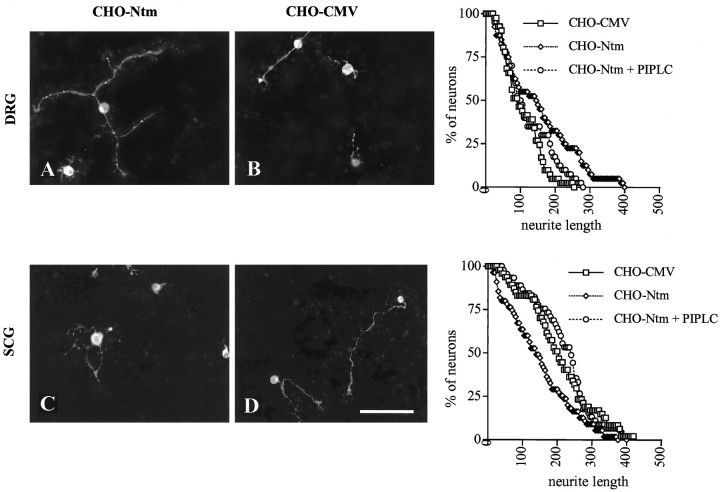
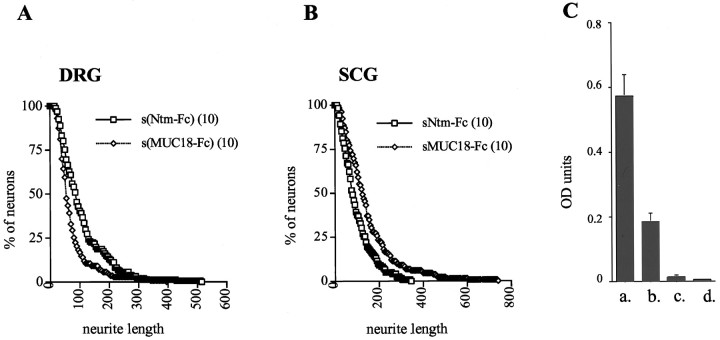
Neurotrimin (Ntm) together with the limbic system-associated membrane protein (LAMP) and the opioid-binding cell adhesion molecule (OBCAM) comprise the IgLON family of neural cell adhesion molecules. These glycosylphosphatidylinositol (GPI)-anchored proteins are expressed in distinct neuronal systems. In the case of Ntm, its expression pattern suggests a role in the development of thalamocortical and pontocerebellar projections (Struyket al., 1995). We have now characterized Ntm's function in cell adhesion and in neurite outgrowth. Cross-linking studies of transfected cells show that Ntm forms noncovalent homodimers and multimers at the cell surface. Ntm mediates homophilic adhesion, as evidenced by the reaggregation of the transfected cells and the specific binding of an Ntm-Fc chimera to these cells. Consistent with these results, Ntm-Fc binds to neurons that express Ntm at high levels, e.g., dorsal root ganglion (DRG) and hippocampal neurons. It does not bind to DRG neurons treated with phosphatidylinositol-specific phospholipase C (PI-PLC) or to sympathetic neurons that do not express Ntm or other members of the IgLON family at significant levels. Ntm promotes the outgrowth of DRG neurons, even after PI-PLC treatment, suggesting that its effects on outgrowth are mediated by heterophilic interactions. Of particular note, both membrane-bound and soluble Ntm inhibit the outgrowth of sympathetic neurons. These results strongly suggest that Ntm, and other members of the IgLON family, regulate the development of neuronal projections via attractive and repulsive mechanisms that are cell type specific and are mediated by homophilic and heterophilic interactions.
Figures
References
-
- Benchimol S, Fuks A, Jothy S, Beauchemin N, Shirota K, Stanners CP. Carcinoembryonic antigen, a human tumor marker functions as an intercellular adhesion molecule. Cell. 1989;57:327–334. - PubMed
-
- Bixby JL, Harris WA. Molecular mechanisms of axon growth and guidance. Annu Rev Cell Biol. 1991;7:117–159. - PubMed
-
- Brewer GJ. Serum-free B27/neurobasal medium supports differentiated growth of neurons from the striatum, substantia nigra, septum, cerebral cortex, cerebellum, and dentate gyrus. J Neurosci Res. 1995;42:674–683. - PubMed
-
- Brümmendorf T, Rathjen FG. Axonal glycoproteins with immunoglobulin- and fibronectin type III-related domains in vertebrates: structural features, binding activities, and signal transduction. J Neurochem. 1993;61:1207–1219. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous