

Control of cyclin D1, p27(Kip1), and cell cycle progression in human capillary endothelial cells by cell shape and cytoskeletal tension
- PMID: 9802905
- PMCID: PMC25607
- DOI: 10.1091/mbc.9.11.3179
Control of cyclin D1, p27(Kip1), and cell cycle progression in human capillary endothelial cells by cell shape and cytoskeletal tension
Abstract
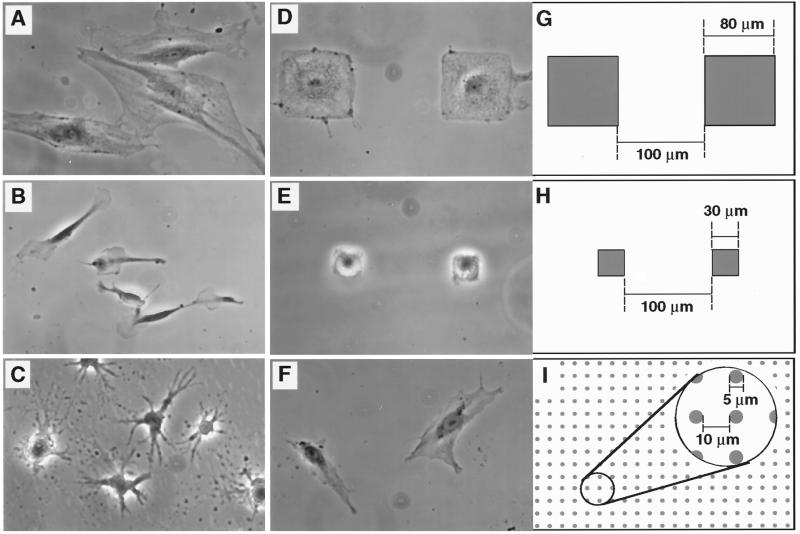
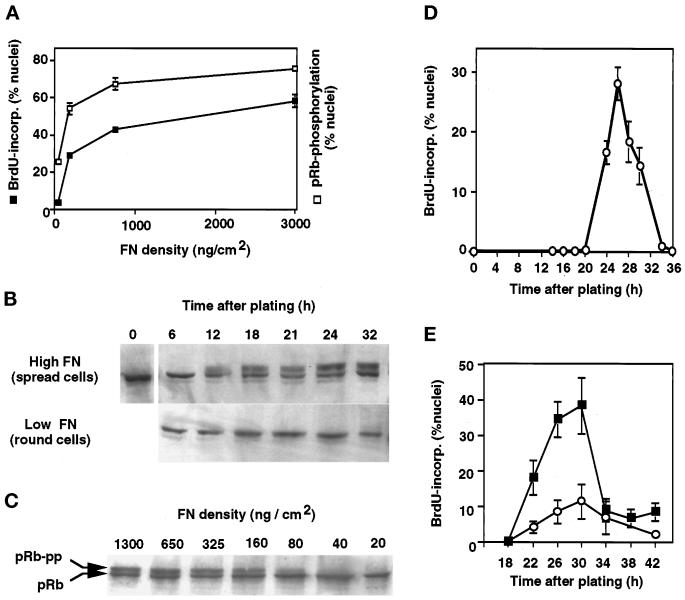
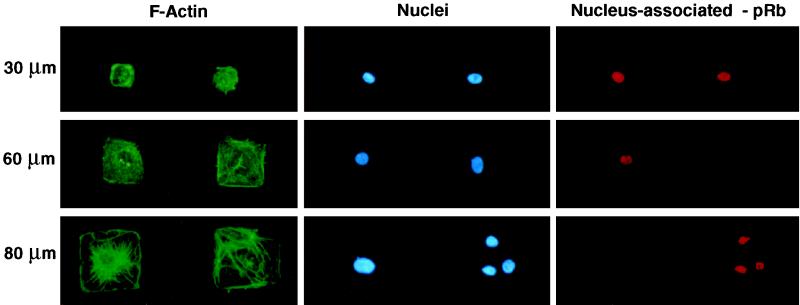
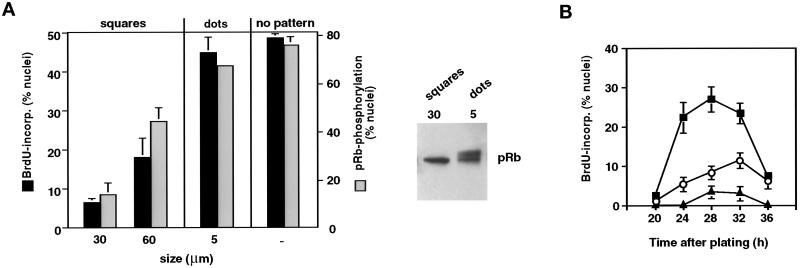
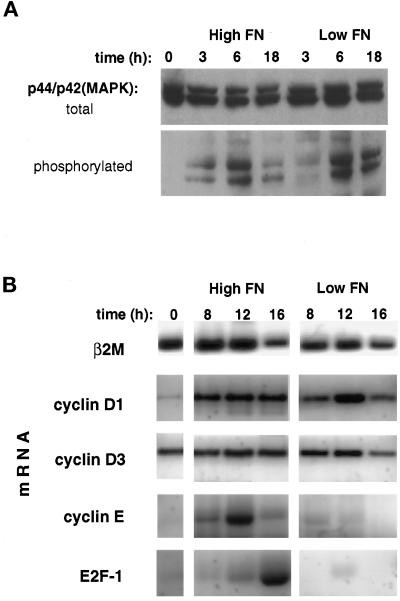
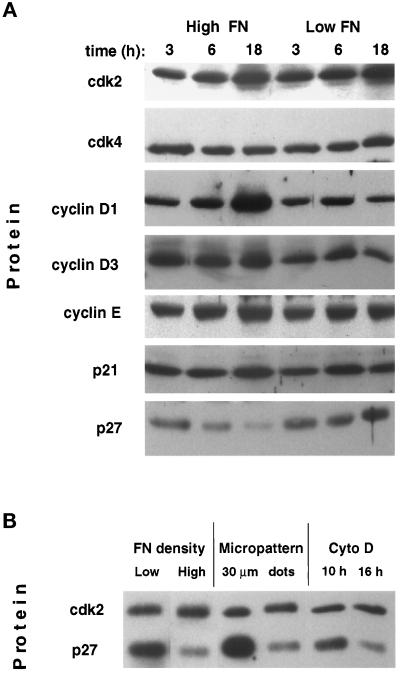
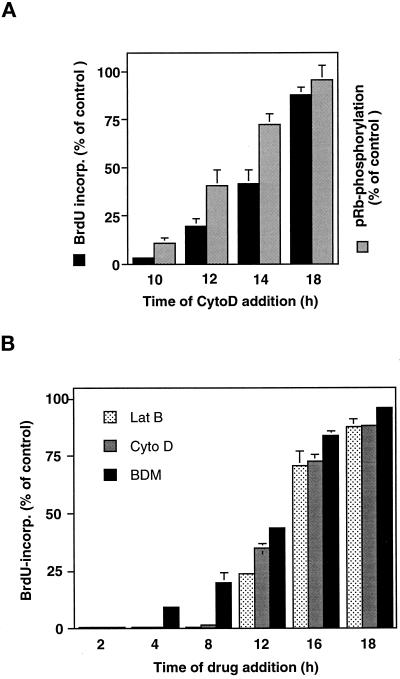
The extracellular matrix (ECM) plays an essential role in the regulation of cell proliferation during angiogenesis. Cell adhesion to ECM is mediated by binding of cell surface integrin receptors, which both activate intracellular signaling cascades and mediate tension-dependent changes in cell shape and cytoskeletal structure. Although the growth control field has focused on early integrin and growth factor signaling events, recent studies suggest that cell shape may play an equally critical role in control of cell cycle progression. Studies were carried out to determine when cell shape exerts its regulatory effects during the cell cycle and to analyze the molecular basis for shape-dependent growth control. The shape of human capillary endothelial cells was controlled by culturing cells on microfabricated substrates containing ECM-coated adhesive islands with defined shape and size on the micrometer scale or on plastic dishes coated with defined ECM molecular coating densities. Cells that were prevented from spreading in medium containing soluble growth factors exhibited normal activation of the mitogen-activated kinase (erk1/erk2) growth signaling pathway. However, in contrast to spread cells, these cells failed to progress through G1 and enter S phase. This shape-dependent block in cell cycle progression correlated with a failure to increase cyclin D1 protein levels, down-regulate the cell cycle inhibitor p27(Kip1), and phosphorylate the retinoblastoma protein in late G1. A similar block in cell cycle progression was induced before this same shape-sensitive restriction point by disrupting the actin network using cytochalasin or by inhibiting cytoskeletal tension generation using an inhibitor of actomyosin interactions. In contrast, neither modifications of cell shape, cytoskeletal structure, nor mechanical tension had any effect on S phase entry when added at later times. These findings demonstrate that although early growth factor and integrin signaling events are required for growth, they alone are not sufficient. Subsequent cell cycle progression and, hence, cell proliferation are controlled by tension-dependent changes in cell shape and cytoskeletal structure that act by subjugating the molecular machinery that regulates the G1/S transition.
Figures
References
-
- Alessandrini A, Chiaur DS, Pagano M. Regulation of the cyclin-dependent kinase inhibitor p27 by degradation and phosphorylation. Leukemia. 1997;11:342–345. - PubMed
-
- Anderson NG, Maller JL, Tonks NK, Sturgill TW. Requirement for integration of signals from two distinct phosphorylation pathways for activation of MAP kinase. Nature. 1990;343:651–653. - PubMed
-
- Assoian RK, Zhu X. Cell anchorage and the cytoskeleton as partners in growth factor dependent cell cycle progression. Curr Opin Cell Biol. 1997;9:93–98. - PubMed
-
- Baldin V, Lukas J, Marcote MJ, Pagano M, Draetta G. Cyclin D1 is a nuclear protein required for cell cycle progression in G1. Genes Dev. 1993;7:812–821. - PubMed
-
- Ben Ze’ev A, Farmer SR, Penman S. Protein synthesis requires cell-surface contact while nuclear events respond to cell shape in anchorage-dependent fibroblasts. Cell. 1980;21:365–372. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
