

Transient alteration of T cell fine specificity by a strong primary stimulus correlates with T cell receptor down-regulation
- PMID: 9808168
- PMCID: PMC2782384
- DOI: 10.1002/(SICI)1521-4141(199810)28:10<2991::AID-IMMU2991>3.0.CO;2-B
Transient alteration of T cell fine specificity by a strong primary stimulus correlates with T cell receptor down-regulation
Abstract
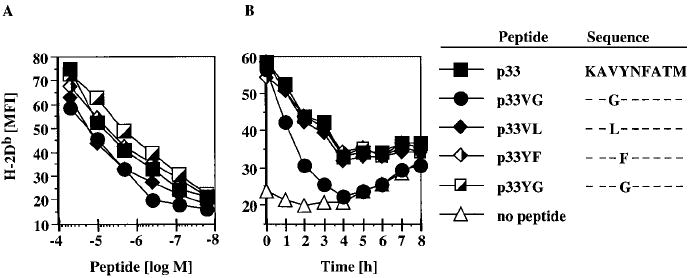
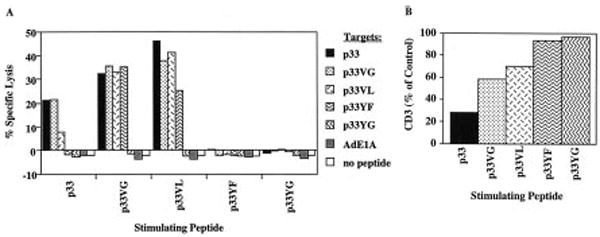
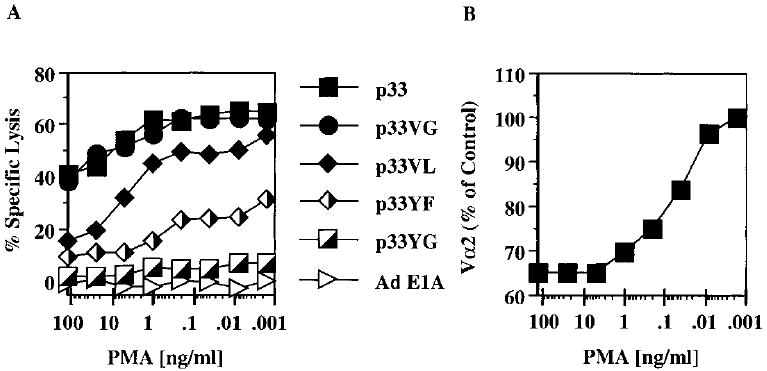
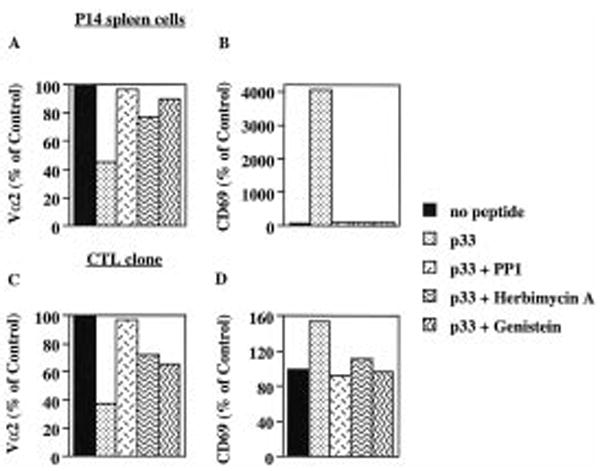
P14 mice expressing a transgenic TCR specific for the lymphocytic choriomeningitis virus glycoprotein p33 epitope were used to study the induction of CTL effector activity by a variety of ligands. Surprisingly, p33 variants which are weaker agonists for the P14 TCR than the wild-type p33 peptide were able to induce more potent effectors with a broader range of cytolytic specificity. Similarly, low concentrations of p33 were more effective than higher concentrations. These results correlated with no or only moderate TCR down-regulation by variants of p33 and low p33 concentrations. This phenotype observed after 18 h of culture was transient as progressive restoration of reactivity was observed at 42 or 66 h in the cultures stimulated with high p33 concentrations and this correlated with recovery of TCR surface levels. TCR down-regulation was blocked by src family kinase inhibitors. These findings indicate that the specificity of a T cell can be fine-tuned by the nature of the primary stimulus correlating with surface TCR level and imply an important role for src family kinases in the differential regulation of surface TCR levels upon TCR engagement by different ligand/MHC complexes.
Figures



References
-
- Sette A, Alexander J, Ruppert J, Snoke K, Franco A, Ishioka G, Grey HM. Antigen analogs/MHC complexes as specific T cell receptor antagonists. Annu Rev Immunol. 1994;12:413–431. - PubMed
-
- Jameson SC, Bevan MJ. T cell receptor antagonists and partial agonists. Immunity. 1995;2:1–11. - PubMed
-
- Sloan-Lancaster J, Allen PM. Altered peptide ligand-induced partial T cell activation: molecular mechanisms and role in T cell biology. Annu Rev Immunol. 1996;14:1–27. - PubMed
-
- Meier U-C, Klenermann P, Griffin P, James W, Köppe B, Larder B, McMichael A, Philipps R. Cytotoxic T lymphocyte lysis inhibited by viable HIV mutants. Science. 1995;270:1360–1362. - PubMed
-
- Bertoletti A, Sette A, Chisari FV, Penna A, Levrero M, De Carli M, Fiaccadori F, Ferrari C. Natural variants of cytotoxic epitopes are T-cell receptor antagonists for antiviral cytotoxic T cells. Nature. 1994;369:407–410. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
