

GAGA mediates the enhancer blocking activity of the eve promoter in the Drosophila embryo
- PMID: 9808619
- PMCID: PMC317233
- DOI: 10.1101/gad.12.21.3325
GAGA mediates the enhancer blocking activity of the eve promoter in the Drosophila embryo
Abstract
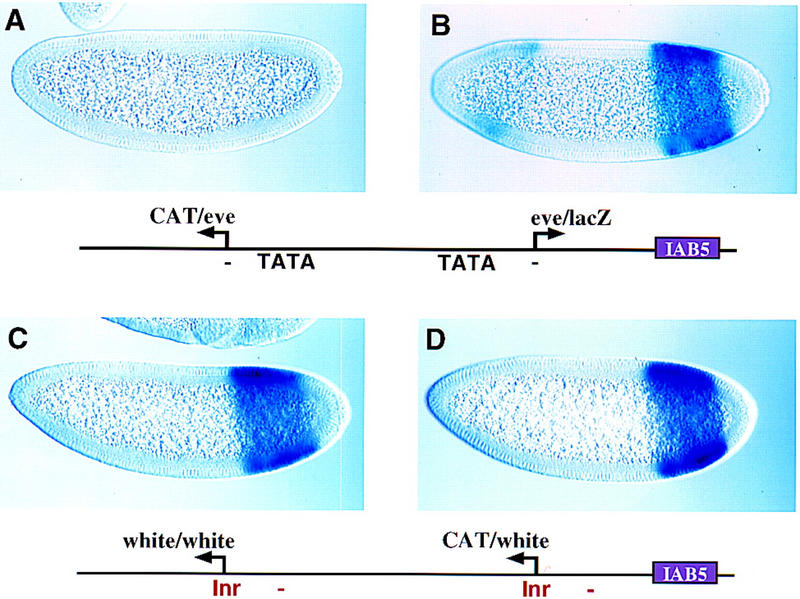
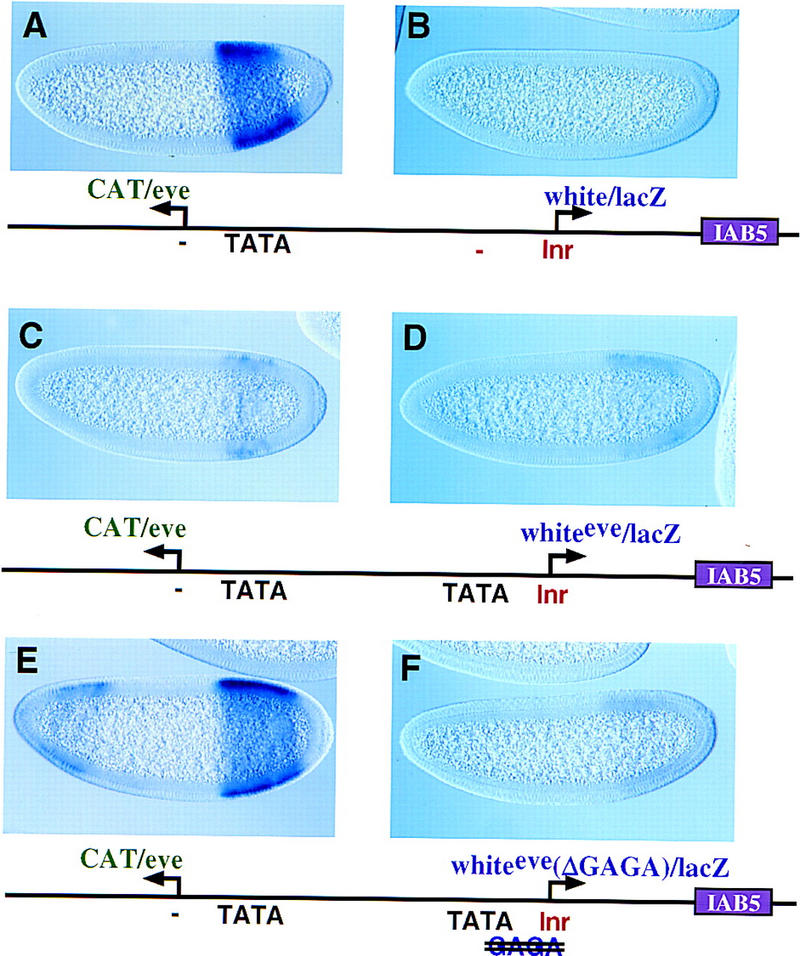
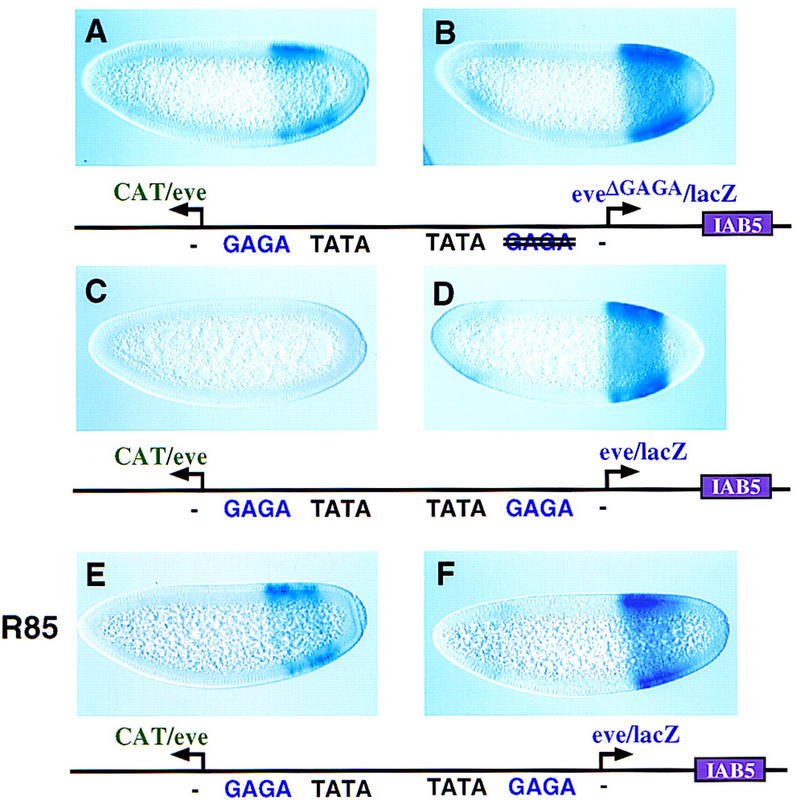
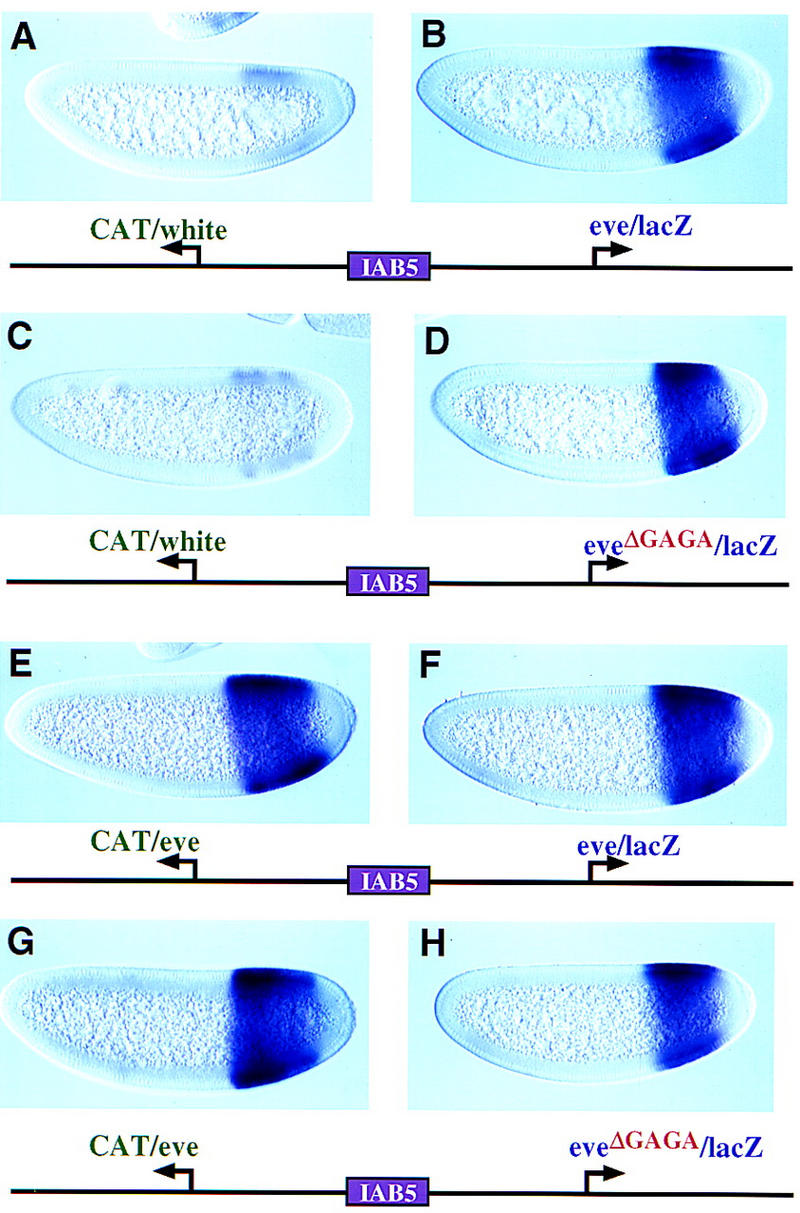
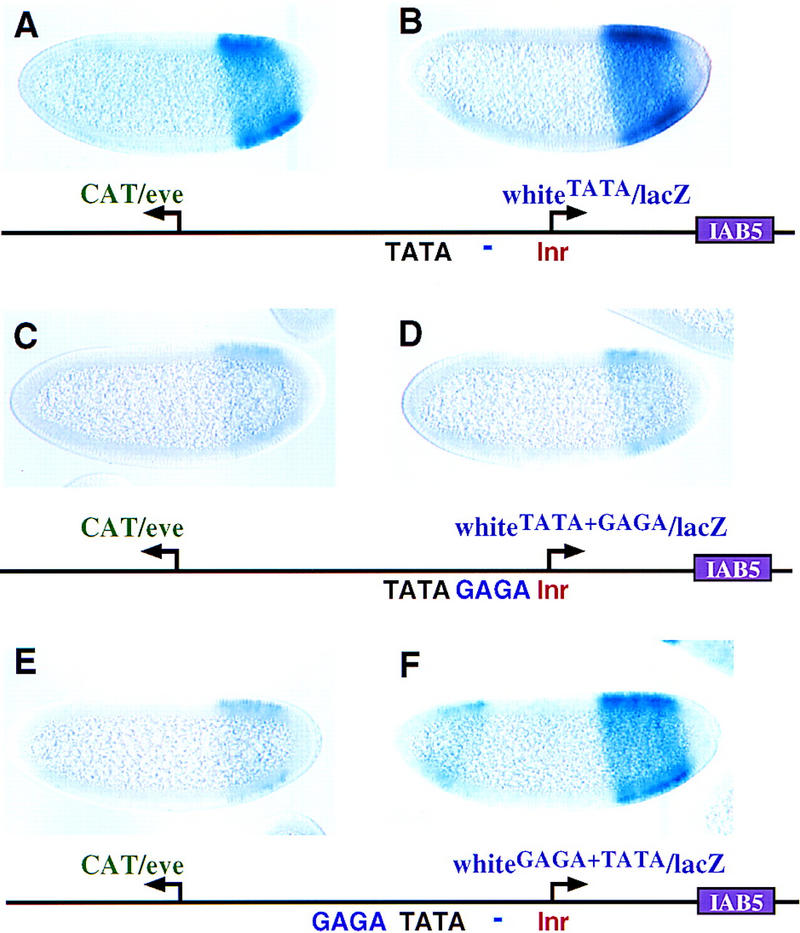
Insulator DNAs and promoter competition regulate enhancer-promoter interactions within complex genetic loci. A transgenic embryo assay was used to obtain evidence that the Drosophila eve promoter possesses an insulator activity that can be uncoupled from the core elements that mediate competition. The eve promoter contains an optimal TATA element and a GAGA sequence. The analysis of various chimeric promoters provides evidence that TATA is essential for promoter competition, whereas GAGA mediates enhancer blocking. The Trithorax-like (Trl) protein interacts with GAGA, and mutations in trl attenuate eve promoter insulator activity. We suggest that Trl-GAGA increases the stability of enhancer-promoter interactions by creating an open chromatin configuration at the core promoter.
Figures
References
-
- Bhat KM, Farkas G, Karch F, Gyurkovics H, Gausz J, Schedl P. The GAGA factor is required in the early Drosophila embryo not only for transcriptional regulation but also for nuclear division. Development. 1996;122:1113–1124. - PubMed
-
- Burke TW, Kadonaga JT. Drosophila TFIID binds to a conserved downsteam basal promoter element that is present in many TAT-box-deficient promoters. Genes & Dev. 1996;10:711–724. - PubMed
-
- Cai H, Levine M. Modulation of enhancer-promoter interactions by insulators in the Drosophila embryo. Nature. 1995;376:533–536. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases