

JNKK1 organizes a MAP kinase module through specific and sequential interactions with upstream and downstream components mediated by its amino-terminal extension
- PMID: 9808624
- PMCID: PMC317229
- DOI: 10.1101/gad.12.21.3369
JNKK1 organizes a MAP kinase module through specific and sequential interactions with upstream and downstream components mediated by its amino-terminal extension
Abstract
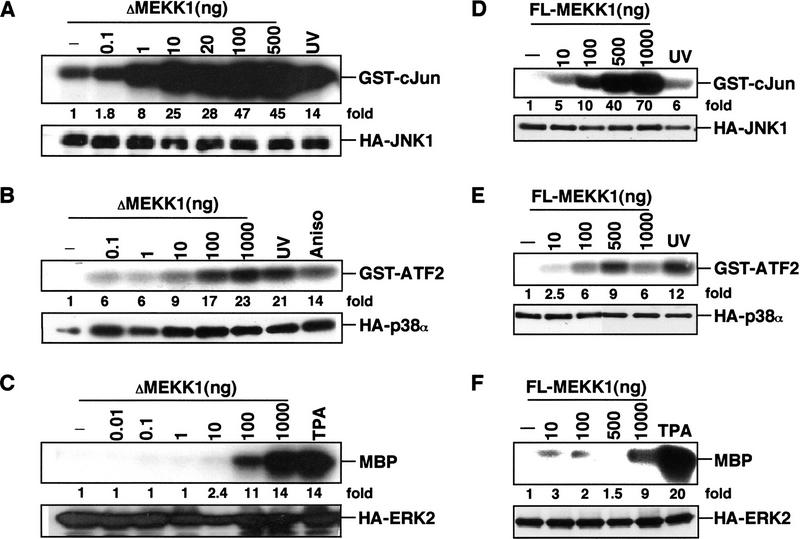
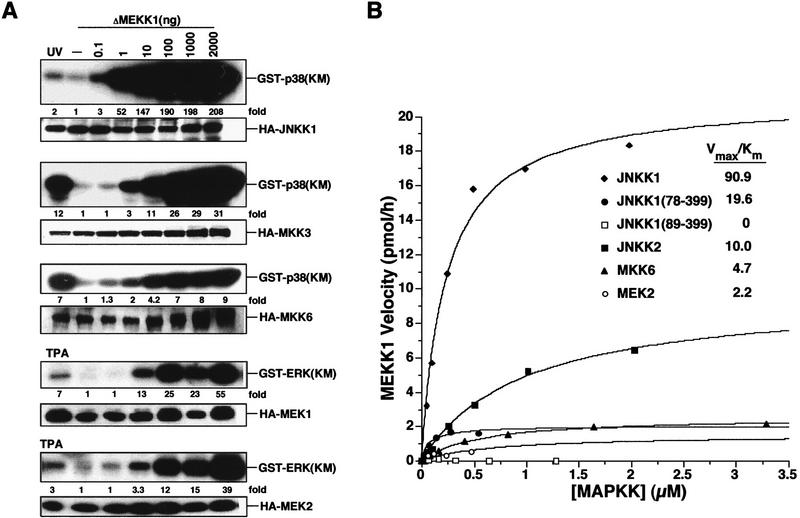
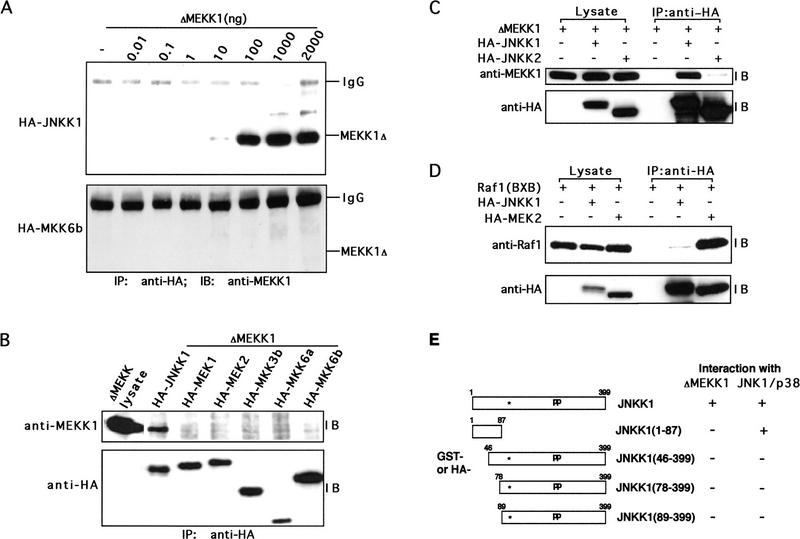
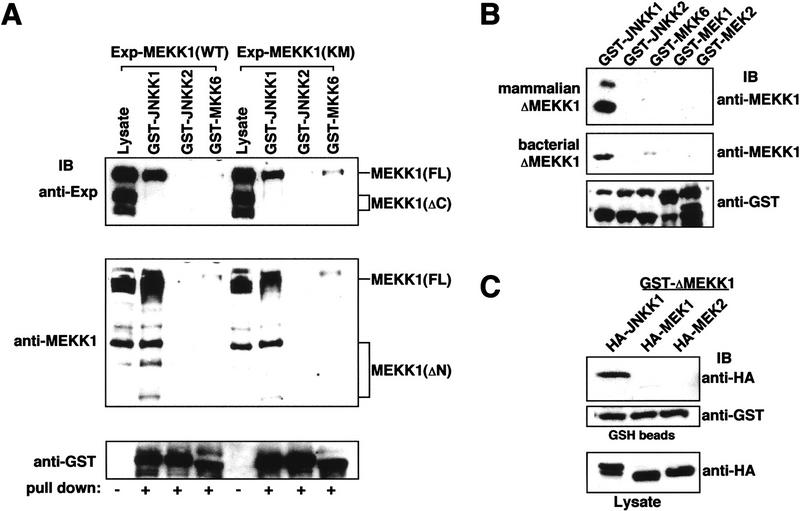
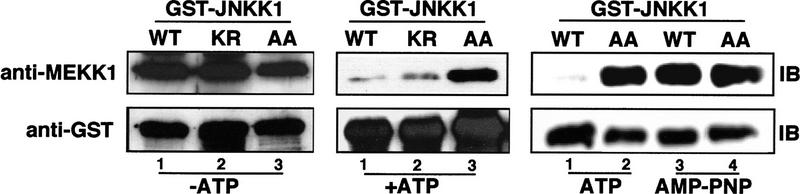
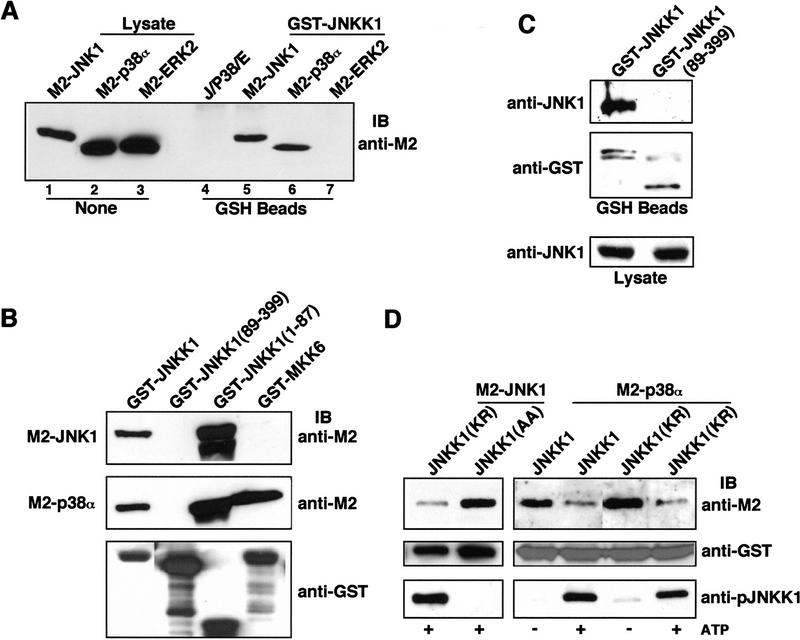
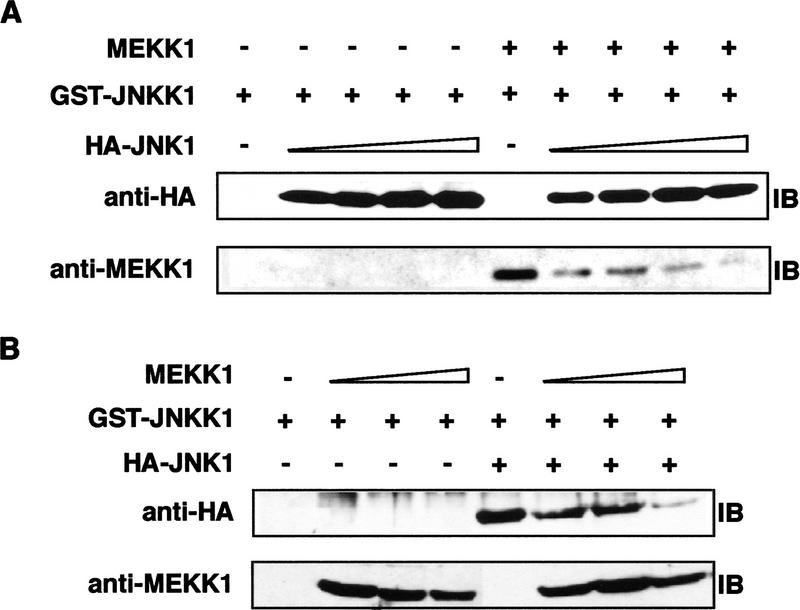
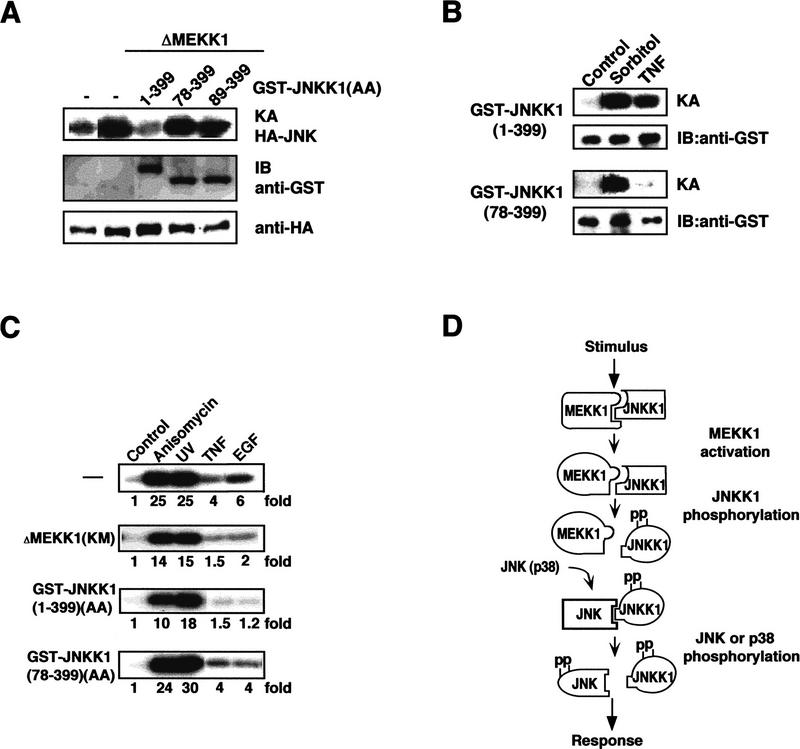
MAP kinase (MAPK) cascades are composed of a MAPK, MAPK kinase (MAPKK), and a MAPKK kinase (MAPKKK). Despite the existence of numerous components and ample opportunities for crosstalk, most MAPKs are specifically and distinctly activated. We investigated the basis for specific activation of the JNK subgroup of MAPKs. The specificity of JNK activation is determined by the MAPKK JNKK1, which interacts with the MAPKKK MEKK1 and JNK through its amino-terminal extension. Inactive JNKK1 mutants can disrupt JNK activation by MEKK1 or tumor necrosis factor (TNF) in intact cells only if they contain an intact amino-terminal extension. Mutations in this region interfere with the ability of JNKK1 to respond to TNF but do not affect its activation by physical stressors. As JNK and MEKK1 compete for binding to JNKK1 and activation of JNKK1 prevents its binding to MEKK1, activation of this module is likely to occur through sequential MEKK1:JNKK1 and JNKK1:JNK interactions. These results underscore a role for the amino-terminal extension of MAPKKs in determination of response specificity.
Figures
References
-
- Avruch J, Zhang X, Kyriakis J. Raf meets ras: Completing the framework of a signal transduction pathway. Trends Biochem Sci. 1994;19:279–283. - PubMed
-
- Bax B, Jhoti H. Protein-protein interactions: putting the pieces together. Curr Biol. 1995;5:1119–1121. - PubMed
-
- Blank JL, Gerwins P, Elliott EM, Sather S, Johnson GL. Molecular cloning of mitogen-activated protein/ERK kinase kinases (MEKK) 2 and 3. J Biol Chem. 1996;274:5361–5368. - PubMed
-
- Brewster JL, de Valoir T, Dwyer ND, Winter E, Gustin MC. An osmosensing signal transduction pathway in yeast. Science. 1993;259:1760–1763. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous