

Bacterial senescence: stasis results in increased and differential oxidation of cytoplasmic proteins leading to developmental induction of the heat shock regulon
- PMID: 9808629
- PMCID: PMC317226
- DOI: 10.1101/gad.12.21.3431
Bacterial senescence: stasis results in increased and differential oxidation of cytoplasmic proteins leading to developmental induction of the heat shock regulon
Abstract
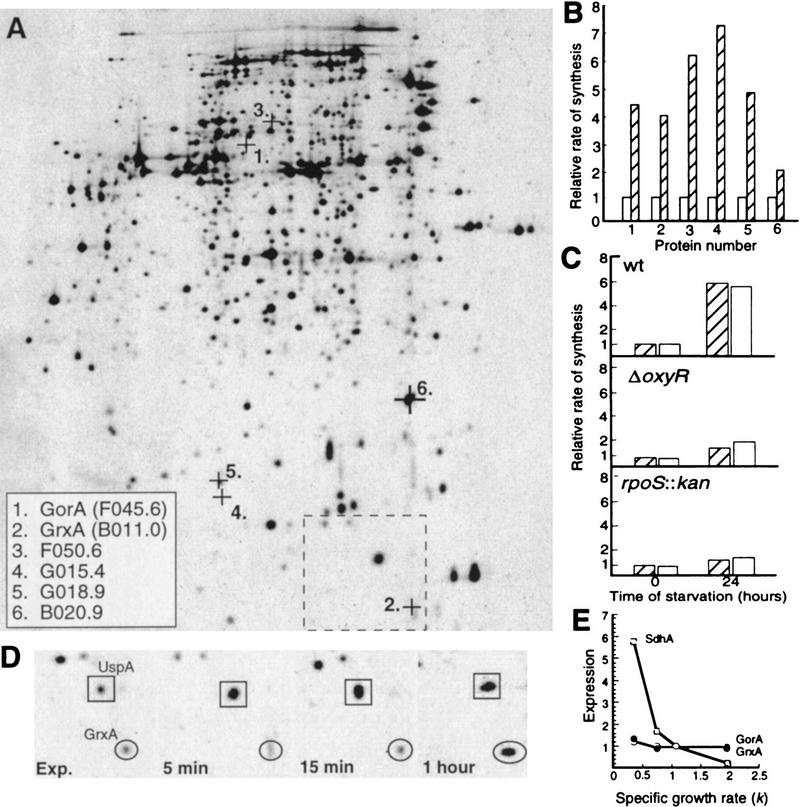
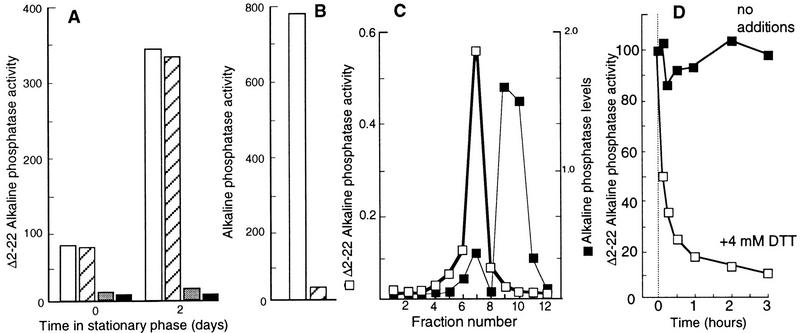
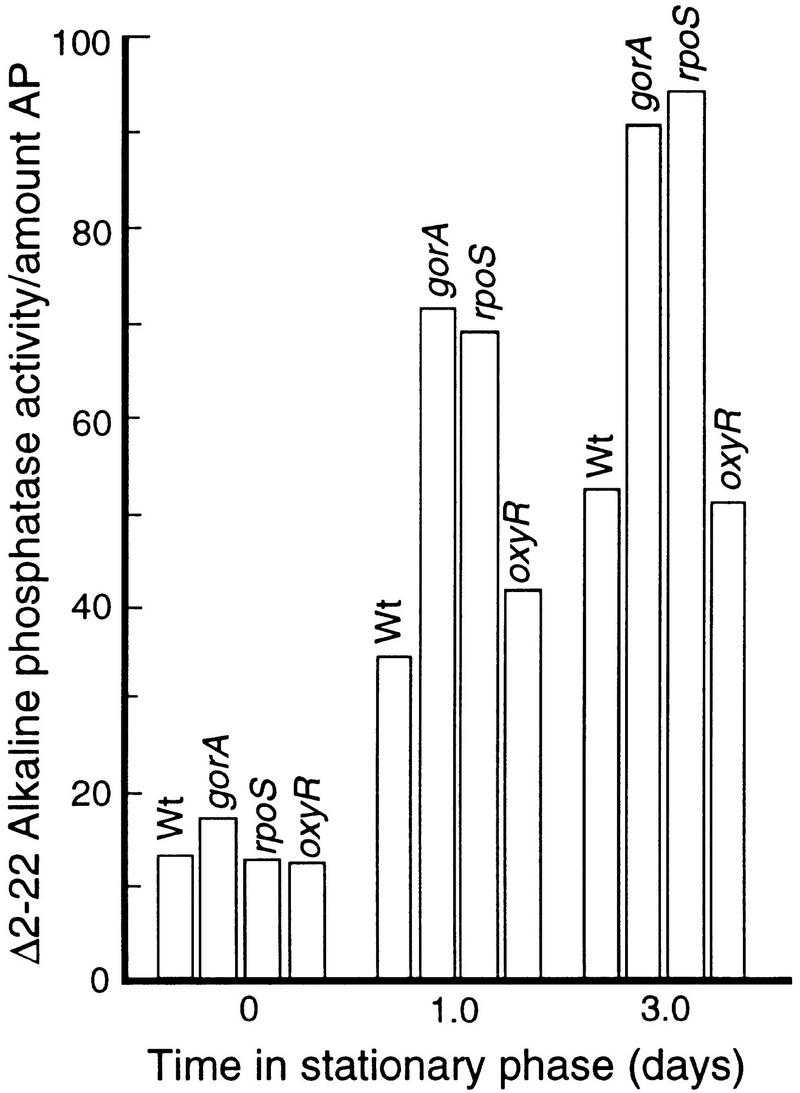
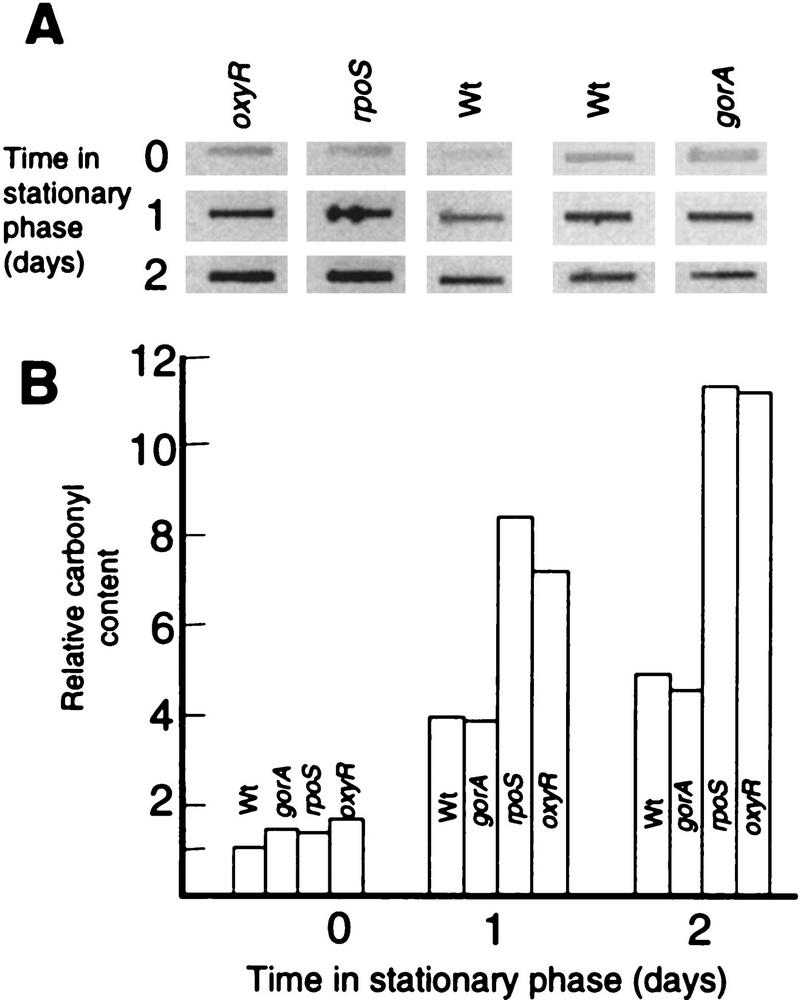
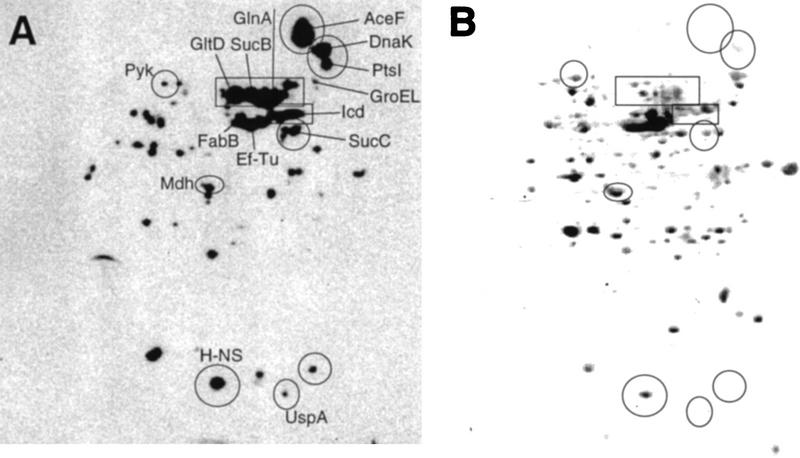
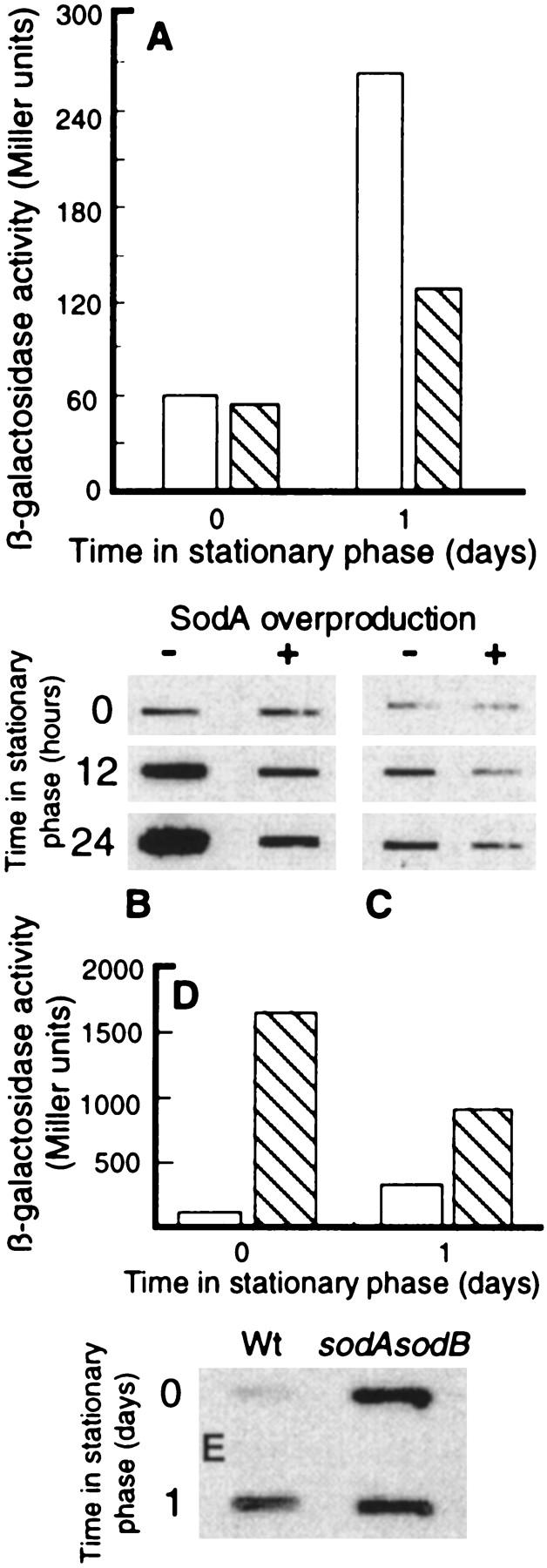
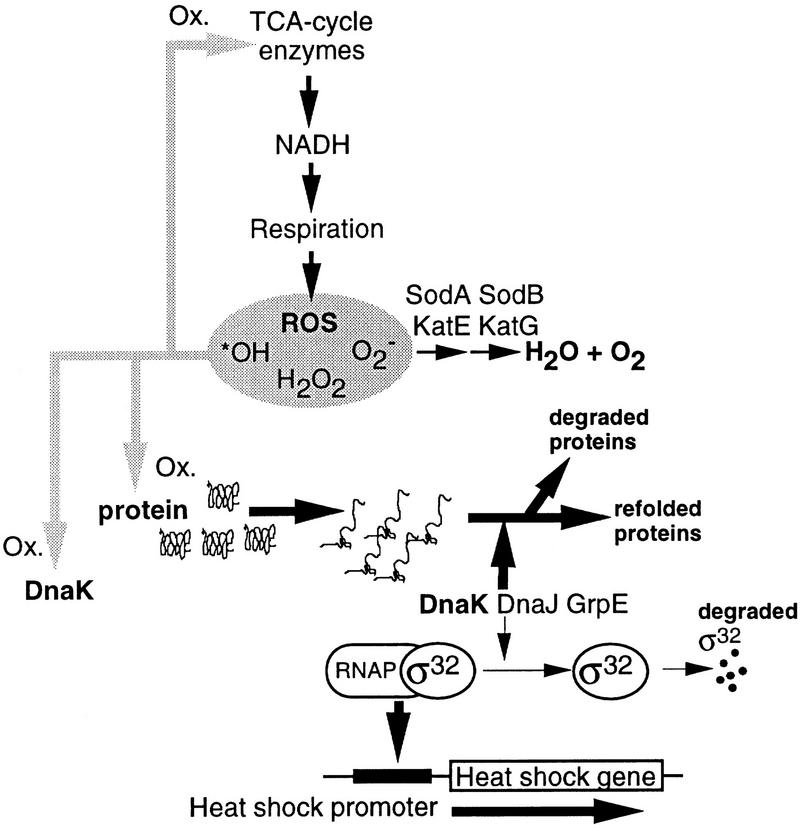
Aging, or senescence, is the progressive deterioration of every bodily function over time. A fundamental question that applies to all life forms, including growth-arrested bacteria, is why growing older by necessity causes organisms to grow more fragile. In this work, we demonstrate that the levels of oxidized proteins is correlated to the age of a stationary-phase Escherichia coli culture; both disulfide bridge formation of a cytoplasmic leader-less alkaline phosphatase and protein carbonyl levels increase during stasis. The stasis-induced increase in protein oxidation is enhanced in cells lacking the global regulators OxyR and sigmas. Some proteins were found to be specifically susceptible to stasis-induced oxidation; notably several TCA cycle enzymes, glutamine synthetase, glutamate synthase, pyruvate kinase, DnaK, and H-NS. Evidence that oxidation of target proteins during stasis serves as the signal for stationary-phase, developmental, induction of the heat shock regulon is presented by demonstrating that this induction is mitigated by overproducing the superoxide dismutase SodA. In addition, cells lacking cytoplasmic superoxide dismutase activity exhibit superinduction of heat shock proteins. The possibility that oxidative sensitivity of TCA cycle enzymes serves as a feedback mechanism down-regulating toxic respiration is discussed.
Figures
References
-
- Albertson NH, Nyström T. Effects of starvation for exogenous carbon on functional mRNA stability and rate of peptide chain elongation in Escherichia coli. FEMS Microbiol Lett. 1994;117:181–187. - PubMed
-
- Almiron M, Link AJ, Furlong D, Kolter R. A novel DNA-binding protein with regulatory and protective roles in starved Escherichia coli. Genes & Dev. 1992;6:2646–2654. - PubMed
-
- Amici A, Levine RL, Tsai L, Stadtman ER. Conversion of amino acid residues in proteins and amino acid homopolymers to carbonyl derivatives by metal-catalyzed oxidation reactions. J Biol Chem. 1989;264:3341–3346. - PubMed
-
- Becker-Hapak M, Eisenstark A. Role of rpoS in the regulation of glutathione oxidoreductase (gor) in Escherichia coli. FEMS Microbiol Lett. 1995;134:39–44. - PubMed
-
- Benov L, Fridovich I. A superoxide dismutase mimic protects sodA sodB Escherichia coli against aerobic heating and stationary-phase death. Arch Biochem Biophys. 1995;322:291–294. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous