

Altered Zn compartmentation in the root symplasm and stimulated Zn absorption into the leaf as mechanisms involved in Zn hyperaccumulation in thlaspi caerulescens
- PMID: 9808732
- PMCID: PMC34798
- DOI: 10.1104/pp.118.3.875
Altered Zn compartmentation in the root symplasm and stimulated Zn absorption into the leaf as mechanisms involved in Zn hyperaccumulation in thlaspi caerulescens
Abstract
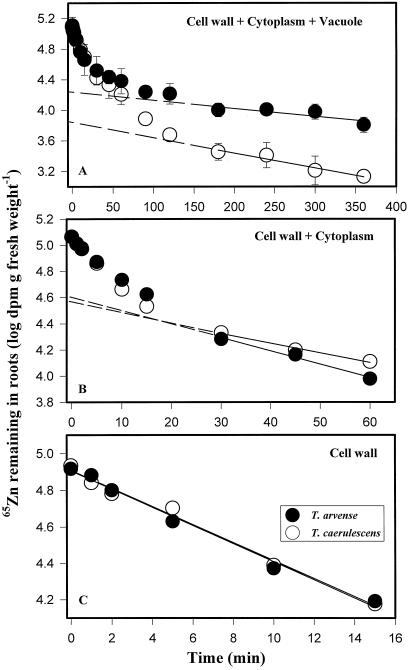
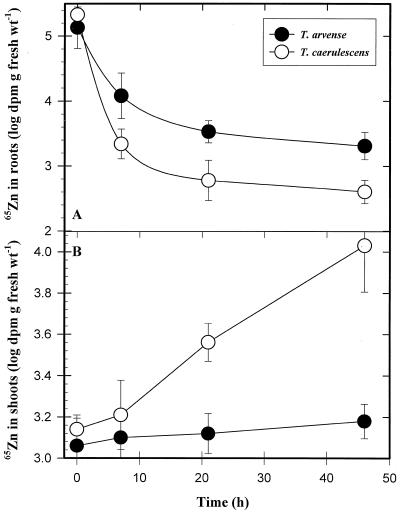
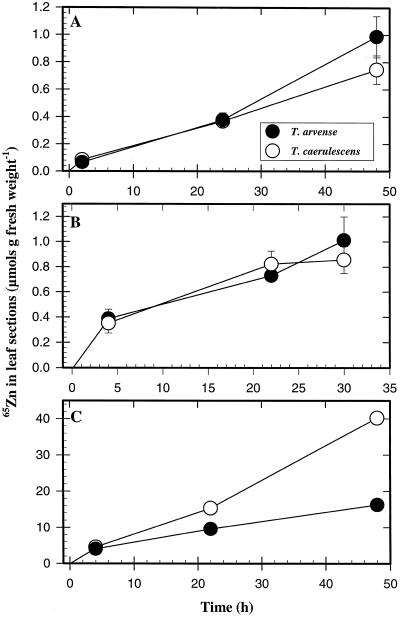
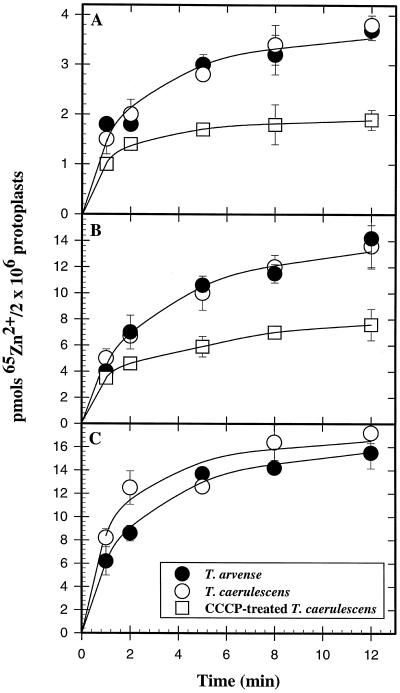
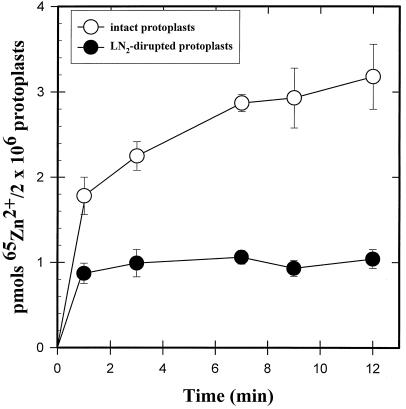
We investigated Zn compartmentation in the root, Zn transport into the xylem, and Zn absorption into leaf cells in Thlaspi caerulescens, a Zn-hyperaccumulator species, and compared them with those of a related nonaccumulator species, Thlaspi arvense. 65Zn-compartmental analysis conducted with roots of the two species indicated that a significant fraction of symplasmic Zn was stored in the root vacuole of T. arvense, and presumably became unavailable for loading into the xylem and subsequent translocation to the shoot. In T. caerulescens, however, a smaller fraction of the absorbed Zn was stored in the root vacuole and was readily transported back into the cytoplasm. We conclude that in T. caerulescens, Zn absorbed by roots is readily available for loading into the xylem. This is supported by analysis of xylem exudate collected from detopped Thlaspi species seedlings. When seedlings of the two species were grown on either low (1 &mgr;M) or high (50 &mgr;M) Zn, xylem sap of T. caerulescens contained approximately 5-fold more Zn than that of T. arvense. This increase was not correlated with a stimulated production of any particular organic or amino acid. The capacity of Thlaspi species cells to absorb 65Zn was studied in leaf sections and leaf protoplasts. At low external Zn levels (10 and 100 &mgr;M), there was no difference in leaf Zn uptake between the two Thlaspi species. However, at 1 mM Zn2+, 2.2-fold more Zn accumulated in leaf sections of T. caerulescens. These findings indicate that altered tonoplast Zn transport in root cells and stimulated Zn uptake in leaf cells play a role in the dramatic Zn hyperaccumulation expressed in T. caerulescens.
Figures
References
-
- Baker AJM, Brooks RR. Terrestrial higher plants which hyperaccumulate metallic elements: a review of their distribution, ecology and phytochemistry. Biorecovery. 1989;1:81–126.
-
- Baker AJM, McGrath SP, Reeves RD, Smith JAC (1998) Metal hyperaccumulator plants: a review of the ecology and physiology of a biological resource for phytoremediation of metal-polluted soils. In N Terry, GS Bañuelos, eds, Phytoremediation. Ann Arbor Press, Ann Arbor, MI (in press)
-
- Baker AJM, Reeves RD, Hajar ASM. Heavy metal accumulation and tolerance in British populations of the metallophyte Thlaspi caerulescens J. & C. Presl (Brassicaceae) New Phytol. 1994;127:61–68. - PubMed
-
- Brown SL, Chaney RL, Angle JS, Baker AJM. Phytoremediation potential of Thlaspi caerulescens and bladder campion for zinc- and cadmium-contaminated soil. J Environ Qual. 1994;23:1151–1157.
-
- Brown SL, Chaney RL, Angle JS, Baker AJM. Zinc and cadmium uptake by hyperaccumulator Thlaspi caerulescens and metal tolerant Silene vulgaris grown on sludge-amended soils. Environ Sci Technol. 1995a;29:1581–1585. - PubMed
LinkOut - more resources
Full Text Sources
Miscellaneous
