

Fitness landscapes for effects of shape on chemotaxis and other behaviors of bacteria
- PMID: 9811657
- PMCID: PMC107673
- DOI: 10.1128/JB.180.22.5978-5983.1998
Fitness landscapes for effects of shape on chemotaxis and other behaviors of bacteria
Abstract
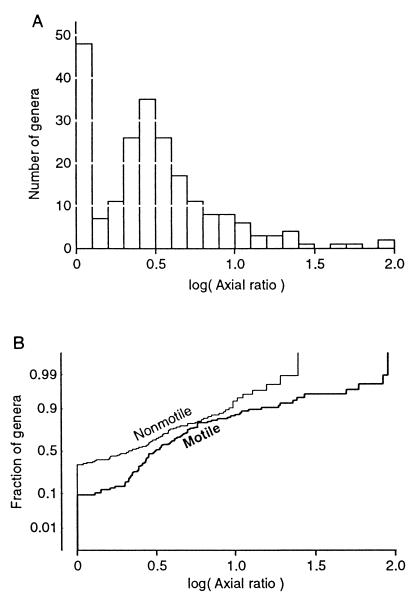
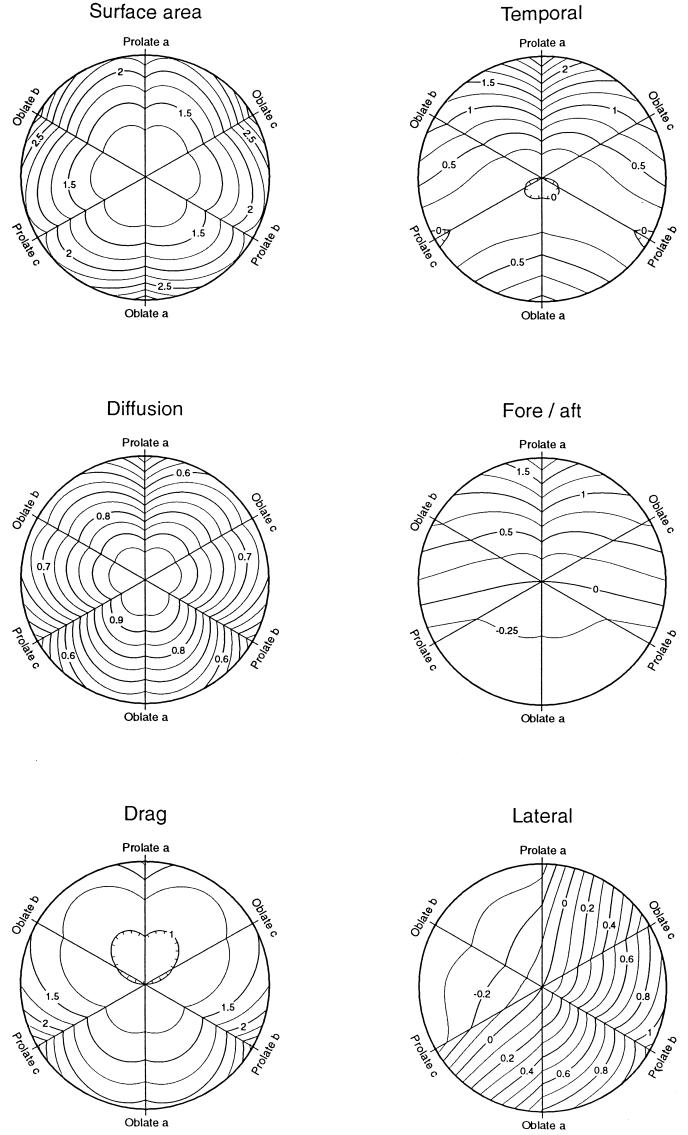
Data on the shapes of 218 genera of free-floating or free-swimming bacteria reveal groupings around spherical shapes and around rod-like shapes of axial ratio about 3. Motile genera are less likely to be spherical and have larger axial ratios than nonmotile genera. The effects of shape on seven possible components of biological fitness were determined, and actual fitness landscapes in phenotype space are presented. Ellipsoidal shapes were used as models, since their hydrodynamic drag coefficients can be rigorously calculated in the world of low Reynolds number, where bacteria live. Comparing various shapes of the same volume, and assuming that departures from spherical have a cost that varies with the minimum radius of curvature, led to the following conclusions. Spherical shapes have the largest random dispersal by Brownian motion. Increased surface area occurs in oblate ellipsoids (disk-like), which rarely occur. Elongation into prolate ellipsoids (rod-like) reduces sinking speed, and this may explain why some nonmotile genera are rod-like. Elongation also favors swimming efficiency (to a limited extent) and the ability to detect stimulus gradients by any of three mechanisms. By far the largest effect (several hundred-fold) is on temporal detection of stimulus gradients, and this explains why rod-like shapes and this mechanism of chemotaxis are common.
Figures
References
-
- Berg H C. Random walks in biology. Princeton, N.J: Princeton University Press; 1993.
-
- Cooper S. Bacterial growth and division. San Diego, Calif: Academic Press; 1991.
-
- Cooper S, Denny M W. A conjecture on the relationship of bacterial shape to motility in rod-shaped bacteria. FEMS Microbiol Lett. 1997;148:227–231.
-
- Cox R G. The motion of long slender bodies in a viscous fluid. Part 1. General theory. J Fluid Mech. 1970;44:791–810.
MeSH terms
LinkOut - more resources
Full Text Sources
