

Interaction of poly(rC) binding protein 2 with the 5' noncoding region of hepatitis A virus RNA and its effects on translation
- PMID: 9811700
- PMCID: PMC110476
- DOI: 10.1128/JVI.72.12.9668-9675.1998
Interaction of poly(rC) binding protein 2 with the 5' noncoding region of hepatitis A virus RNA and its effects on translation
Abstract
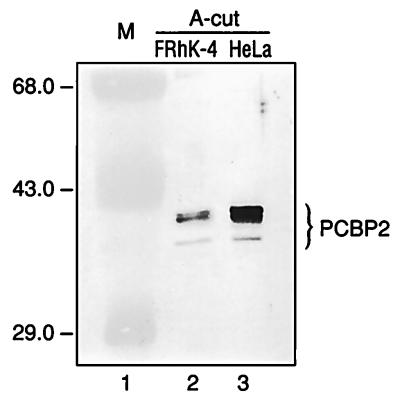
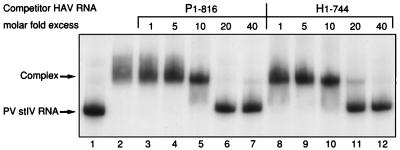
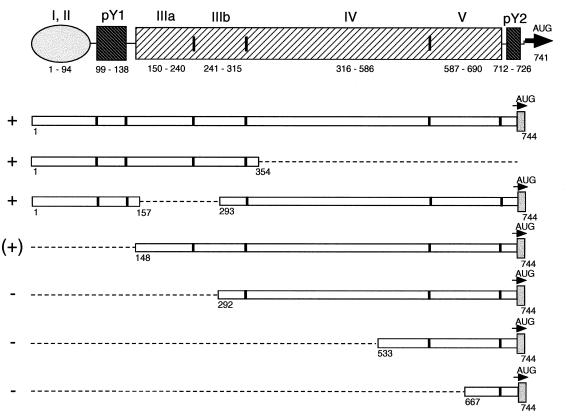
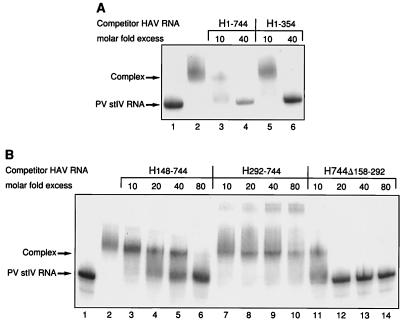
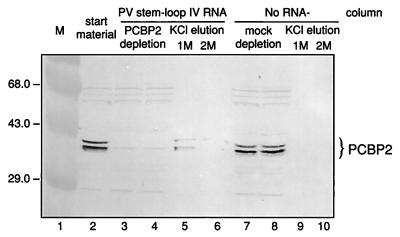
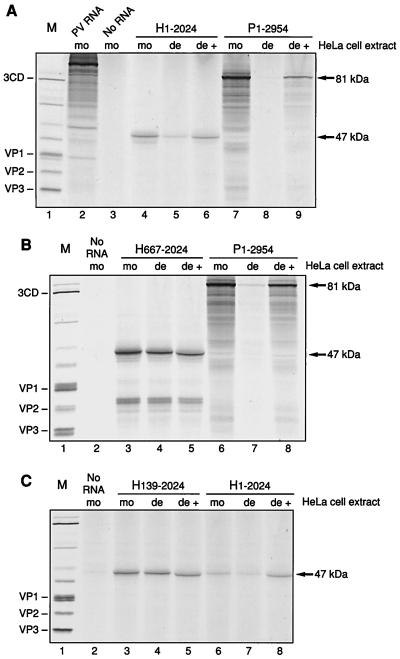
Utilization of internal ribosome entry segment (IRES) structures in the 5' noncoding region (5'NCR) of picornavirus RNAs for initiation of translation requires a number of host cell factors whose distribution may vary in different cells and whose requirement may vary for different picornaviruses. We have examined the requirement of the cellular protein poly(rC) binding protein 2 (PCBP2) for hepatitis A virus (HAV) RNA translation. PCBP2 has recently been identified as a factor required for translation and replication of poliovirus (PV) RNA. PCBP2 was shown to be present in FRhK-4 cells, which are permissive for growth of HAV, as it is in HeLa cells, which support translation of HAV RNA but which have not been reported to host replication of the virus. Competition RNA mobility shift assays showed that the 5'NCR of HAV RNA competed for binding of PCBP2 with a probe representing stem-loop IV of the PV 5'NCR. The binding site on HAV RNA was mapped to nucleotides 1 to 157, which includes a pyrimidine-rich sequence. HeLa cell extracts that had been depleted of PCBP2 by passage over a PV stem-loop IV RNA affinity column supported only low levels of HAV RNA translation. Translation activity was restored upon addition of recombinant PCBP2 to the depleted extract. Removal of the 5'-terminal 138 nucleotides of the HAV RNA, or removal of the entire IRES, eliminated the dependence of HAV RNA translation on PCBP2.
Figures
Similar articles
-
Differential utilization of poly(rC) binding protein 2 in translation directed by picornavirus IRES elements.RNA. 1999 Dec;5(12):1570-85. doi: 10.1017/s1355838299991483. RNA. 1999. PMID: 10606268 Free PMC article.
-
Distinct poly(rC) binding protein KH domain determinants for poliovirus translation initiation and viral RNA replication.J Virol. 2002 Dec;76(23):12008-22. doi: 10.1128/jvi.76.23.12008-12022.2002. J Virol. 2002. PMID: 12414943 Free PMC article.
-
The linker domain of poly(rC) binding protein 2 is a major determinant in poliovirus cap-independent translation.Virology. 2008 Sep 1;378(2):243-53. doi: 10.1016/j.virol.2008.05.007. Epub 2008 Jul 25. Virology. 2008. PMID: 18656221 Free PMC article.
-
Advances and Breakthroughs in IRES-Directed Translation and Replication of Picornaviruses.mBio. 2023 Apr 25;14(2):e0035823. doi: 10.1128/mbio.00358-23. Epub 2023 Mar 20. mBio. 2023. PMID: 36939331 Free PMC article. Review.
-
Mechanistic intersections between picornavirus translation and RNA replication.Adv Virus Res. 2011;80:1-24. doi: 10.1016/B978-0-12-385987-7.00001-4. Adv Virus Res. 2011. PMID: 21762819 Review.
Cited by
-
Giardiavirus internal ribosome entry site has an apparently unique mechanism of initiating translation.PLoS One. 2009 Oct 14;4(10):e7435. doi: 10.1371/journal.pone.0007435. PLoS One. 2009. PMID: 19826476 Free PMC article.
-
Translation of Hepatitis A Virus IRES Is Upregulated by a Hepatic Cell-Specific Factor.Front Genet. 2018 Aug 10;9:307. doi: 10.3389/fgene.2018.00307. eCollection 2018. Front Genet. 2018. PMID: 30147706 Free PMC article.
-
Structure and RNA binding of the third KH domain of poly(C)-binding protein 1.Nucleic Acids Res. 2005 Feb 24;33(4):1213-21. doi: 10.1093/nar/gki265. Print 2005. Nucleic Acids Res. 2005. PMID: 15731341 Free PMC article.
-
Direct-acting Antivirals and Host-targeting Agents against the Hepatitis A Virus.J Clin Transl Hepatol. 2015 Sep 28;3(3):205-10. doi: 10.14218/JCTH.2015.00016. Epub 2015 Sep 15. J Clin Transl Hepatol. 2015. PMID: 26623267 Free PMC article. Review.
-
Heterogeneous nuclear ribonucleoprotein C modulates translation of c-myc mRNA in a cell cycle phase-dependent manner.Mol Cell Biol. 2003 Jan;23(2):708-20. doi: 10.1128/MCB.23.2.708-720.2003. Mol Cell Biol. 2003. PMID: 12509468 Free PMC article.
References
-
- Blyn, L. B., and J. Graff. Unpublished data.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
