

Adeno-associated virus type 2-mediated gene transfer: role of epidermal growth factor receptor protein tyrosine kinase in transgene expression
- PMID: 9811719
- PMCID: PMC110495
- DOI: 10.1128/JVI.72.12.9835-9843.1998
Adeno-associated virus type 2-mediated gene transfer: role of epidermal growth factor receptor protein tyrosine kinase in transgene expression
Abstract
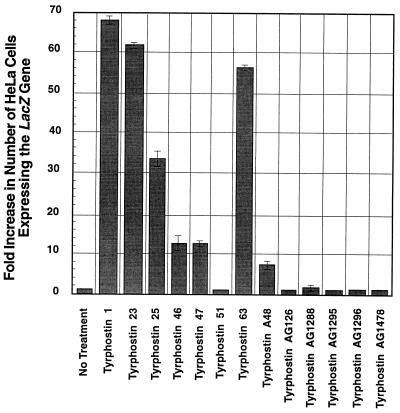
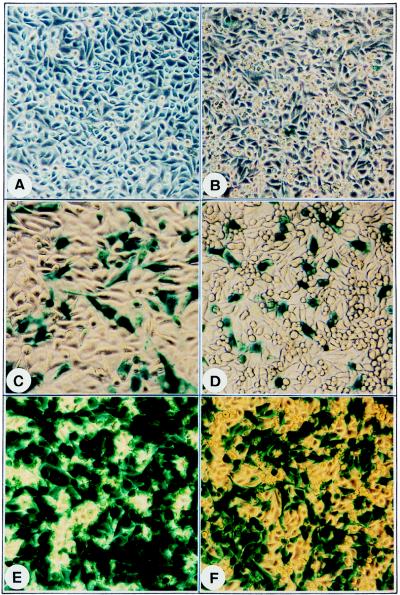
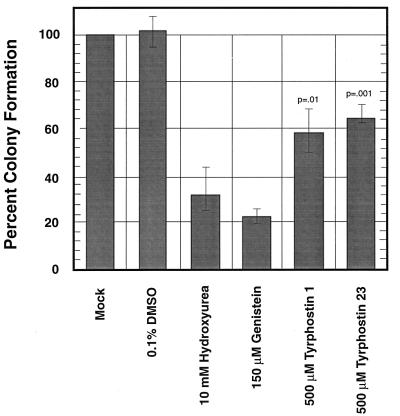
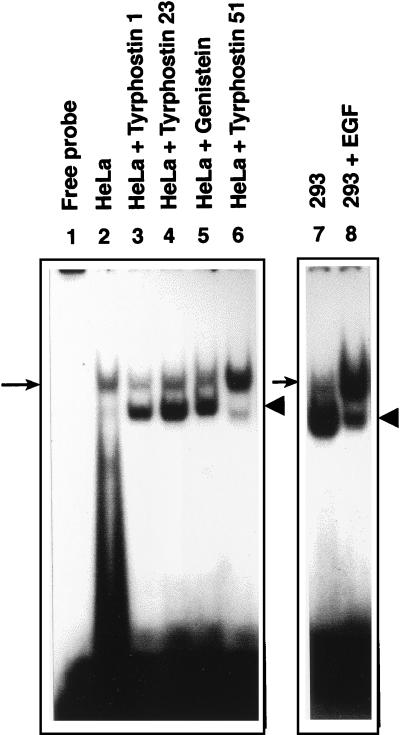
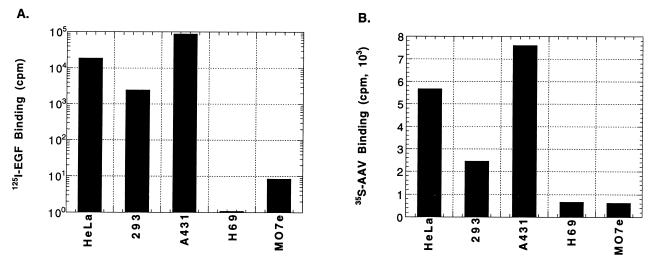
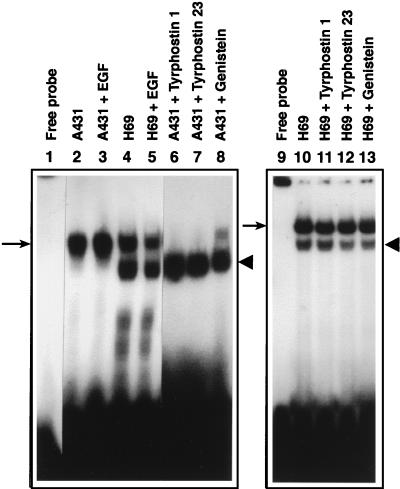
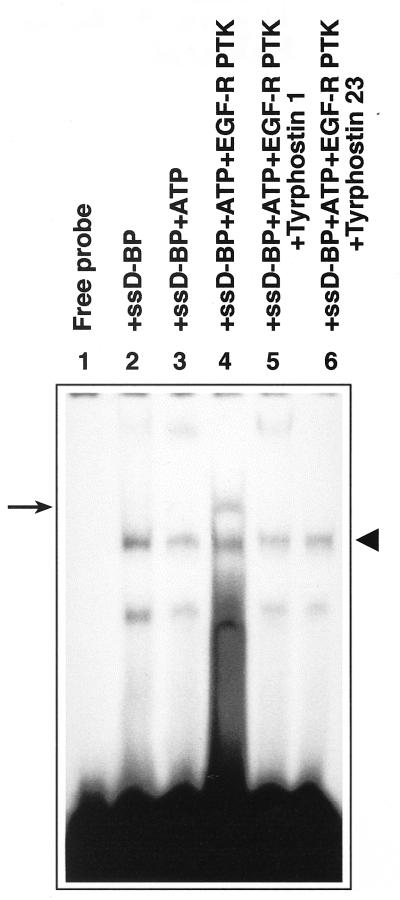
Adeno-associated virus type 2 (AAV), a single-stranded, DNA-containing, nonpathogenic human parvovirus, has gained attention as a potentially useful vector for human gene therapy. However, the transduction efficiency of AAV vectors varies greatly in different cells and tissues in vitro and in vivo. We have recently documented that a cellular tyrosine phosphoprotein, designated the single-stranded D-sequence-binding protein (ssD-BP), plays an important role in AAV-mediated transgene expression (K. Y. Qing et al., Proc. Natl. Acad. Sci. USA 94:10879-10884, 1997) and that a strong correlation exists between the phosphorylation state of the ssD-BP and AAV transduction efficiency in vitro as well as in vivo (K. Y. Qing et al., J. Virol. 72:1593-1599, 1998). In this report, we document that treatment of cells with specific inhibitors of the epidermal growth factor receptor protein tyrosine kinase (EGF-R PTK) activity, such as tyrphostin, leads to significant augmentation of AAV transduction efficiency, and phosphorylation of the ssD-BP is mediated by the EGF-R PTK. Treatment of cells with EGF results in phosphorylation of the ssD-BP, whereas treatment with tyrphostin causes dephosphorylation of the ssD-BP and consequently leads to increased expression of the transgene. Furthermore, AAV transduction efficiency inversely correlates with expression of the EGF-R in different cell types, and stable transfection of the EGF-R cDNA causes phosphorylation of the ssD-BP, leading to significant inhibition in AAV-mediated transgene expression which can be overcome by the tyrphostin treatment. These data suggest that the PTK activity of the EGF-R is a crucial determinant in the life cycle of AAV and that further studies on the interaction between the EGF-R and the ssD-BP may yield new insights not only into its role in the host cell but also in the successful use of AAV vectors in human gene therapy.
Figures

References
-
- Akiyama T, Ishida J, Nakagawa S, Ogawara H, Watanabe S, Itoh N, Shibuya M, Fukami Y. Genistein, a specific inhibitor of tyrosine-specific protein kinases. J Biol Chem. 1987;262:5592–5595. - PubMed
-
- Alexander I E, Russell D W, Spence A M, Miller A D. Effects of gamma irradiation on the transduction of dividing and nondividing cells in brain and muscle of rats by recombinant adeno-associated virus vectors. Hum Gene Ther. 1996;7:841–850. - PubMed
-
- Barnes S, Peterson G T. Biochemical targets of the isoflavone genistein in tumor cell lines. Soc Exp Biol Med. 1995;280:103–109. - PubMed
-
- Berns K I, Bohenzky R A. Adeno-associated viruses: an update. Adv Virus Res. 1987;32:243–307. - PubMed
-
- Berns K I, Giraud C. Biology of adeno-associated virus. Curr Top Microbiol Immunol. 1996;218:1–23. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
