

Genetic analysis of the rat leukemia virus: influence of viral sequences in transduction of the c-ras proto-oncogene and expression of its transforming activity
- PMID: 9811727
- PMCID: PMC110503
- DOI: 10.1128/JVI.72.12.9906-9917.1998
Genetic analysis of the rat leukemia virus: influence of viral sequences in transduction of the c-ras proto-oncogene and expression of its transforming activity
Abstract
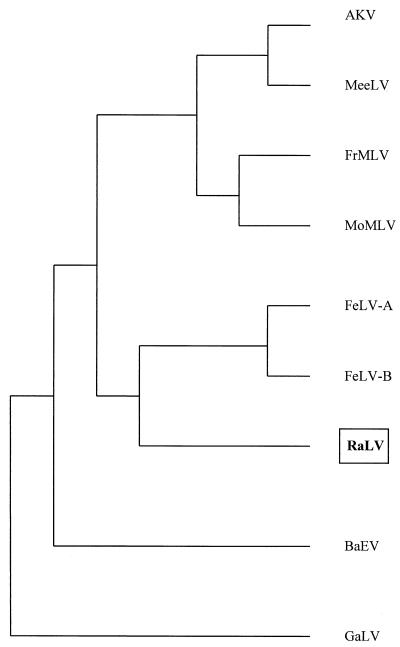
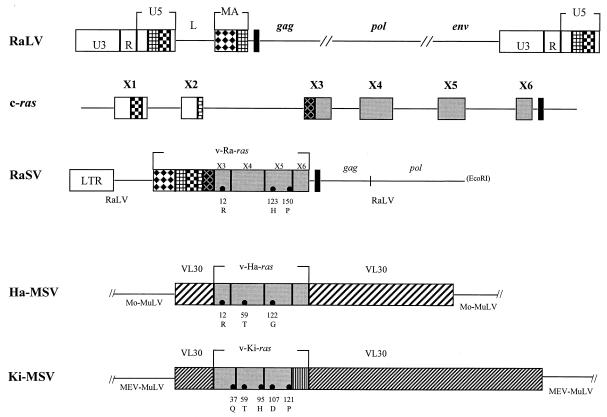
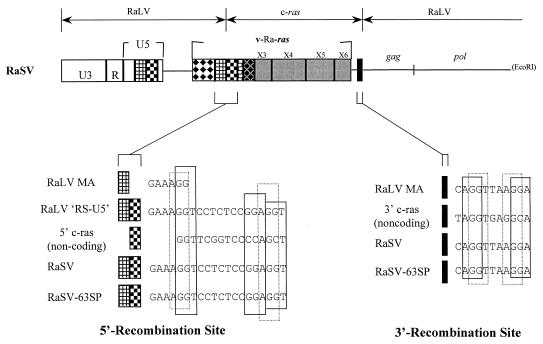
The rat leukemia virus (RaLV) is an endogenous retrovirus that is spontaneously released by Sprague-Dawley rat embryo cells. The overall structure of the RaLV genome resembles that of other simple, replication-competent retroviruses, but the sequence of the long terminal repeats (LTR) is unique and unrelated to the known retroviruses. Phylogenetically, the RaLV genome appears to be more closely related to the feline leukemia virus group of retroviruses than to the murine leukemia virus group. A remarkable feature of RaLV is that it is capable of transducing a ras proto-oncogene from rat tumor cells in the form of an acutely transforming virus, designated the Rasheed strain of the rat sarcoma virus (RaSV). With the exception of the c-ras sequence, the genomes of both RaLV and RaSV are collinear. The RaSV-encoded oncogene v-Ra-ras expresses a fusion protein with a molecular mass of 29 kDa, and it exhibits a unique structure that has not been described previously for any known virus. The 5' end of this gene is derived from sequences encoding RaLV matrix followed by 20 bp derived from the U5 region of the RaLV LTR (RS-U5 element) which is joined at its 3' end to sequences derived from all six (coding and noncoding) exons of the c-ras proto-oncogene at the 3' end. This recombinational event represents a novel mechanism among the acutely transforming viruses that have been studied.
Figures




References
-
- Biessmann H, Mason J M. Telomeric repeat sequences. Chromosoma. 1994;103:154–161. - PubMed
-
- Buss J E, Solski P A, Schaeffer J P, MacDonald M J, Der C J. Activation of the cellular proto-oncogene product p21Ras by addition of a myristylation signal. Science. 1989;243:1600–1603. - PubMed
-
- Capon D J, Chen E Y, Levinson A D, Seeburg P H, Goeddel D V. Complete nucleotide sequences of the T24 human bladder carcinoma oncogene and its normal homologue. Nature. 1983;302:33–37. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
