

KATP channel inhibition by ATP requires distinct functional domains of the cytoplasmic C terminus of the pore-forming subunit
- PMID: 9811907
- PMCID: PMC24984
- DOI: 10.1073/pnas.95.23.13953
KATP channel inhibition by ATP requires distinct functional domains of the cytoplasmic C terminus of the pore-forming subunit
Abstract
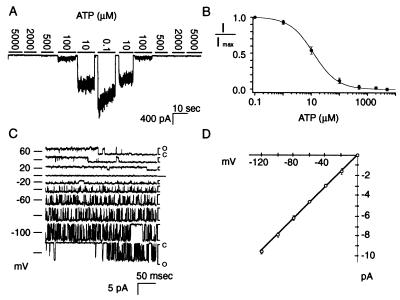
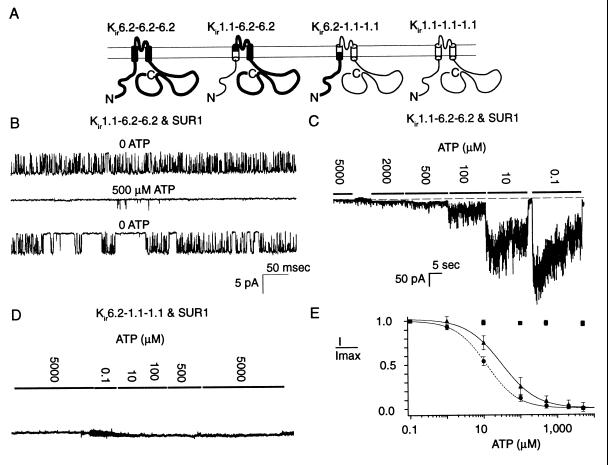
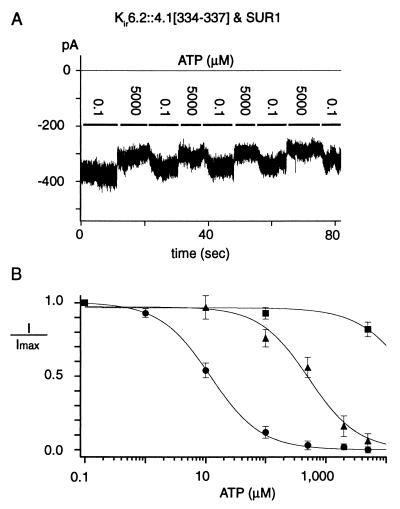
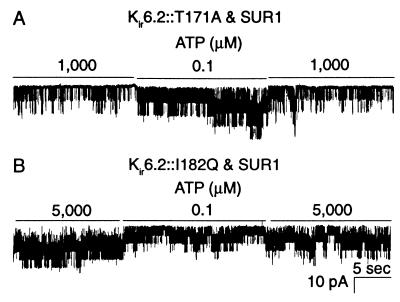
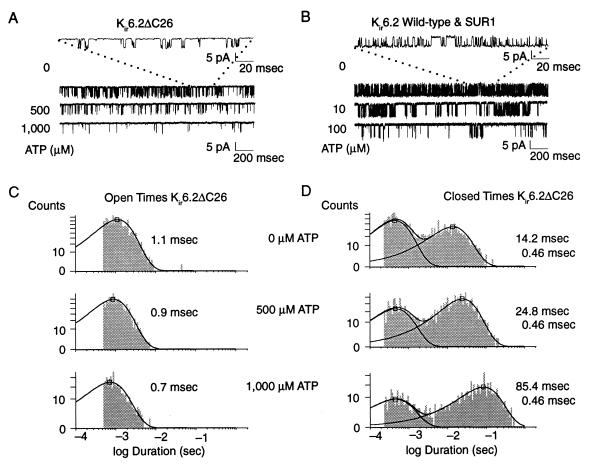
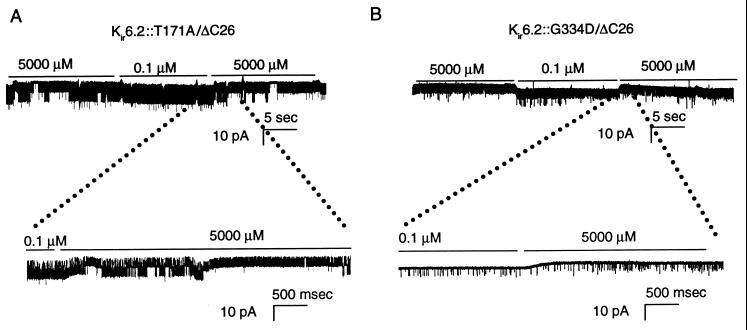
ATP-sensitive potassium ("KATP") channels are rapidly inhibited by intracellular ATP. This inhibition plays a crucial role in the coupling of electrical activity to energy metabolism in a variety of cells. The KATP channel is formed from four each of a sulfonylurea receptor (SUR) regulatory subunit and an inwardly rectifying potassium (Kir6.2) pore-forming subunit. We used systematic chimeric and point mutagenesis, combined with patch-clamp recording, to investigate the molecular basis of ATP-dependent inhibition gating of mouse pancreatic beta cell KATP channels expressed in Xenopus oocytes. We identified distinct functional domains of the presumed cytoplasmic C-terminal segment of the Kir6.2 subunit that play an important role in this inhibition. Our results suggest that one domain is associated with inhibitory ATP binding and another with gate closure.
Figures
References
-
- Inagaki N, Gonoi T, Clement J P, IV, Namba N, Inazawa J, Gonzalez G, Aguilar-Bryan L, Seino S, Bryan J. Science. 1995;270:1166–1170. - PubMed
-
- Clement J P, IV, Kunjilwar K, Gonzales G, Schwanstecher M, Panten U, Aguilar-Bryan L, Bryan J. Neuron. 1997;18:827–838. - PubMed
-
- Inagaki N, Gonoi T, Seino S. FEBS Lett. 1997;409:232–236. - PubMed
-
- Aguilar-Bryan L, Nichols C G, Wechsler S W, Clement J P T, IV, Boyd A E, Gonzalez G, Herrera-Sosa H, Nguy K, Bryan J, Nelson D A. Science. 1995;268:423–426. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
