

ZAP-70 association with T cell receptor zeta (TCRzeta): fluorescence imaging of dynamic changes upon cellular stimulation
- PMID: 9813084
- PMCID: PMC2148150
- DOI: 10.1083/jcb.143.3.613
ZAP-70 association with T cell receptor zeta (TCRzeta): fluorescence imaging of dynamic changes upon cellular stimulation
Abstract
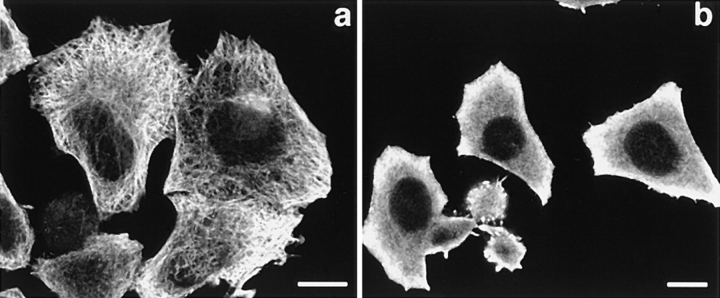
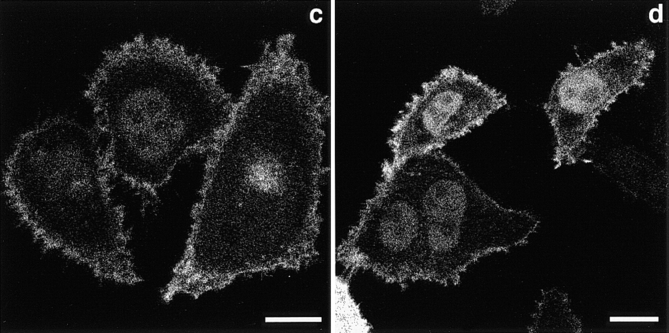
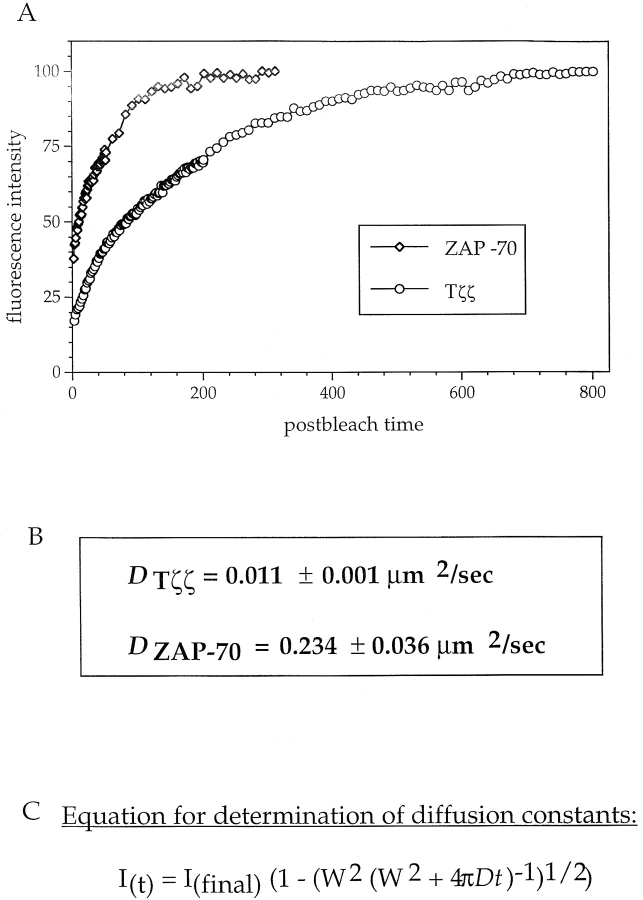
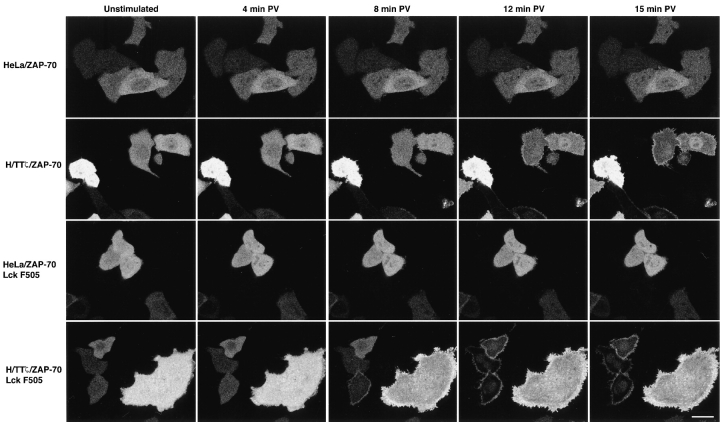
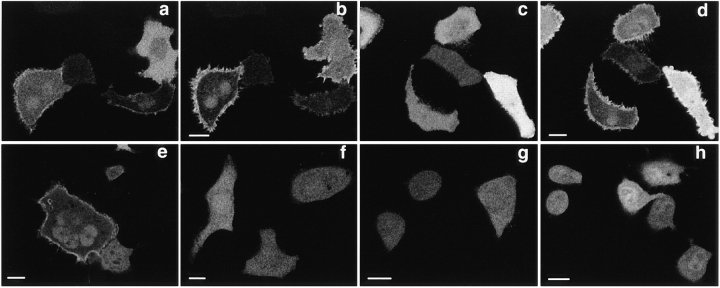
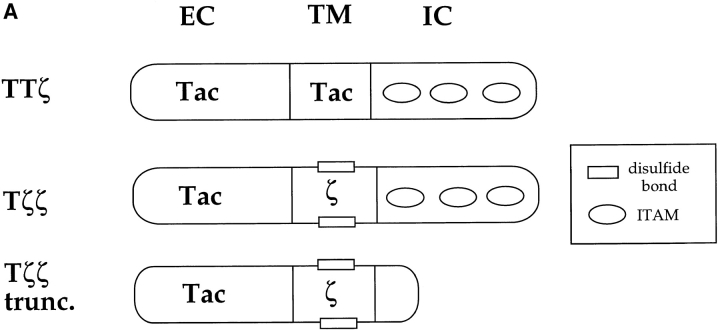
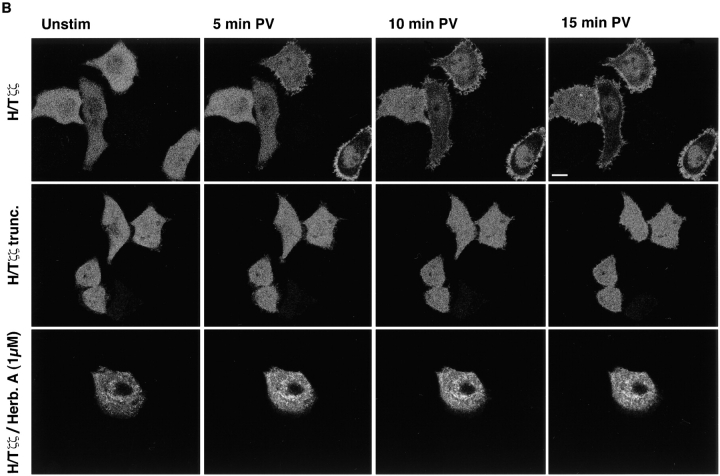
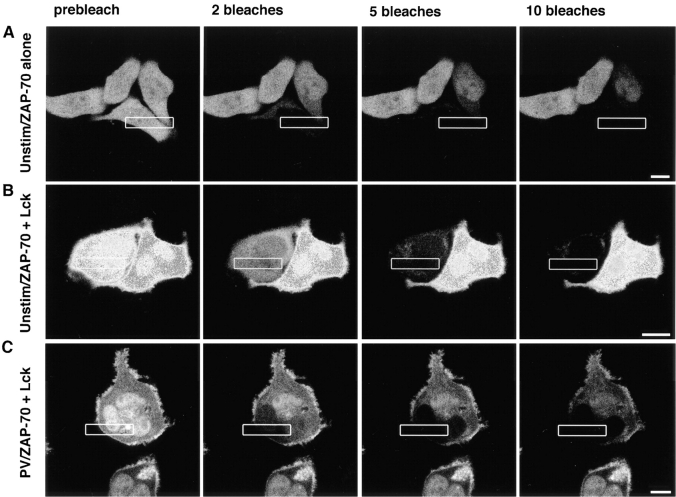
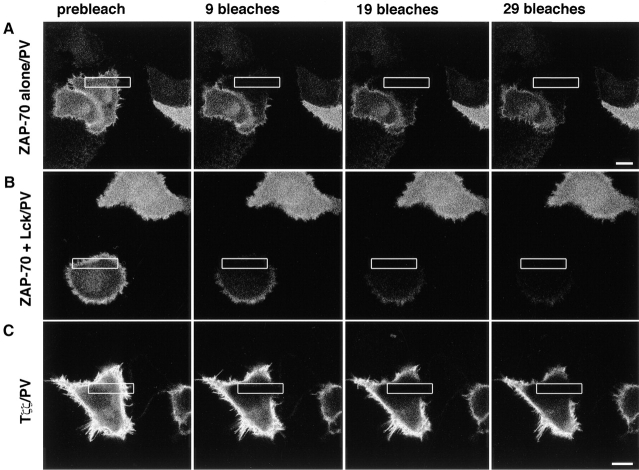
The nonreceptor protein tyrosine kinase ZAP-70 is a critical enzyme required for successful T lymphocyte activation. After antigenic stimulation, ZAP-70 rapidly associates with T cell receptor (TCR) subunits. The kinetics of its translocation to the cell surface, the properties of its specific interaction with the TCRzeta chain expressed as a chimeric protein (TTzeta and Tzetazeta), and its mobility in different intracellular compartments were studied in individual live HeLa cells, using ZAP-70 and Tzetazeta fused to green fluorescent protein (ZAP-70 GFP and Tzetazeta-GFP, respectively). Time-lapse imaging using confocal microscopy indicated that the activation-induced redistribution of ZAP-70 to the plasma membrane, after a delayed onset, is of long duration. The presence of the TCRzeta chain is critical for the redistribution, which is enhanced when an active form of the protein tyrosine kinase Lck is coexpressed. Binding specificity to TTzeta was indicated using mutant ZAP-70 GFPs and a truncated zeta chimera. Photobleaching techniques revealed that ZAP-70 GFP has decreased mobility at the plasma membrane, in contrast to its rapid mobility in the cytosol and nucleus. Tzetazeta- GFP is relatively immobile, while peripherally located ZAP-70 in stimulated cells is less mobile than cytosolic ZAP-70 in unstimulated cells, a phenotype confirmed by determining the respective diffusion constants. Examination of the specific molecular association of signaling proteins using these approaches has provided new insights into the TCRzeta-ZAP-70 interaction and will be a powerful tool for continuing studies of lymphocyte activation.
Figures
References
-
- Arpaia E, Shahar M, Dadi H, Cohen A, Roifman CM. Defective T cell receptor signaling and CD8+thymocyte selection in humans lacking ZAP-70 kinase. Cell. 1994;76:947–958. - PubMed
-
- Babbitt BP, Allen PM, Matsueda G, Haber E, Unanue ER. Binding of immunogenic peptides to Ia histocompatibility molecules. Nature. 1985;317:359–361. - PubMed
-
- Bentley GA, Mariuzza RA. The structure of the T cell antigen receptor. Annu Rev Immunol. 1996;14:563–590. - PubMed
-
- Cantrell D. T cell antigen receptor signal transduction pathways. Annu Rev Immunol. 1996;14:259–274. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
