

Polyunsaturated fatty acids inhibit T cell signal transduction by modification of detergent-insoluble membrane domains
- PMID: 9813086
- PMCID: PMC2148157
- DOI: 10.1083/jcb.143.3.637
Polyunsaturated fatty acids inhibit T cell signal transduction by modification of detergent-insoluble membrane domains
Abstract
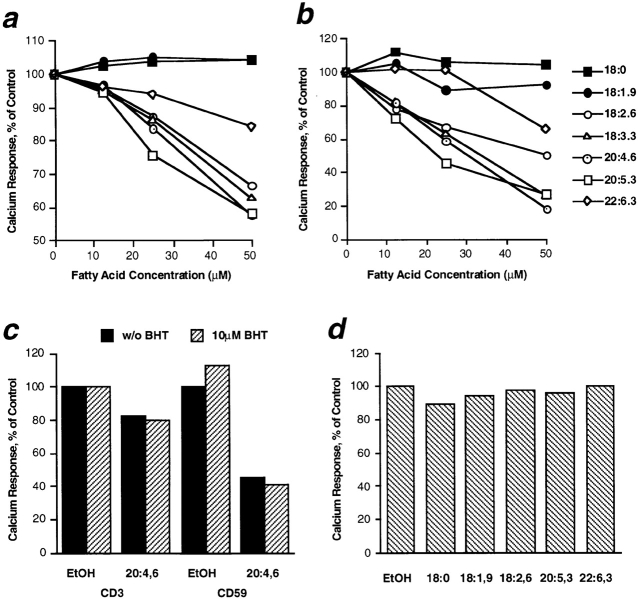
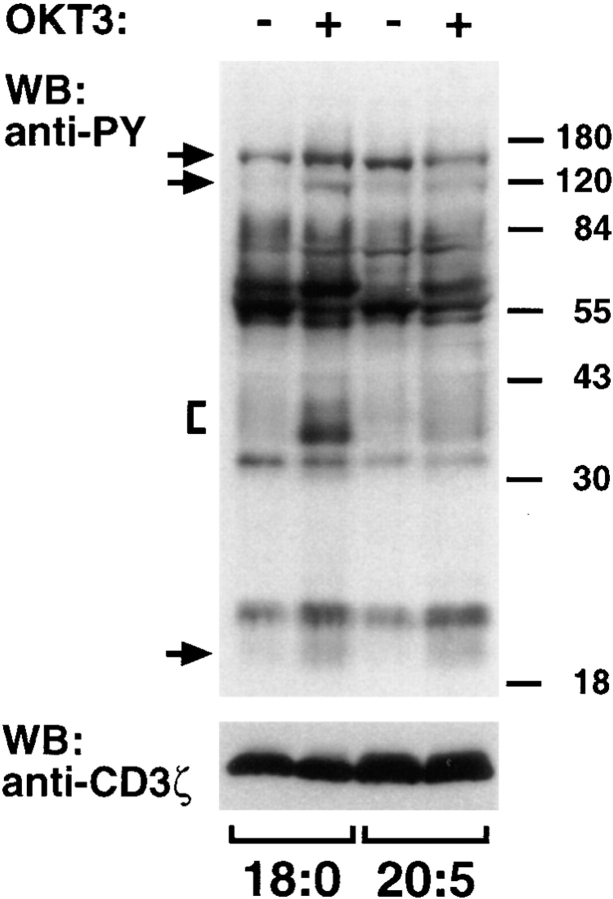
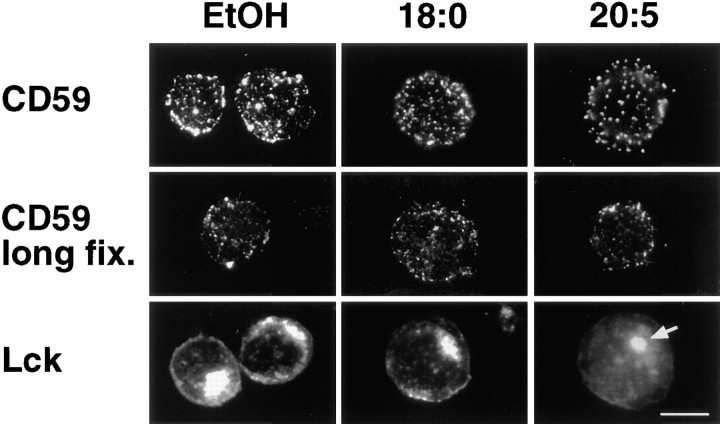
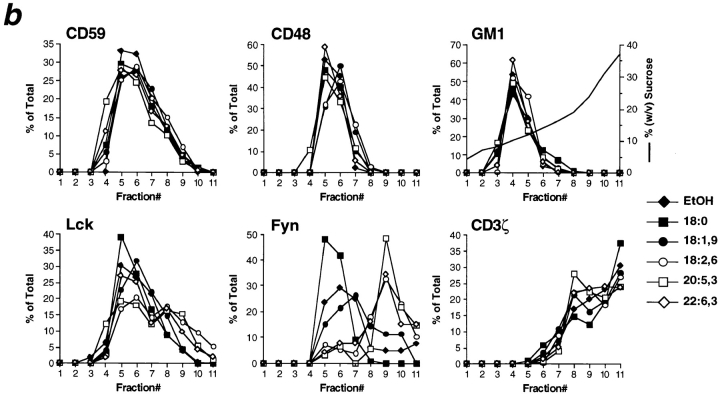
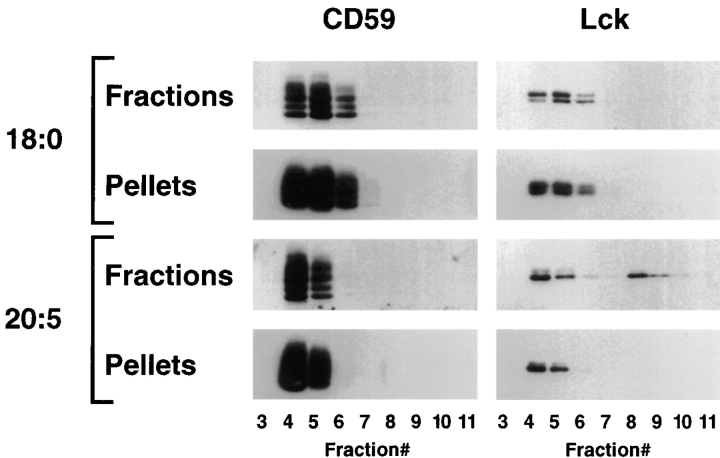
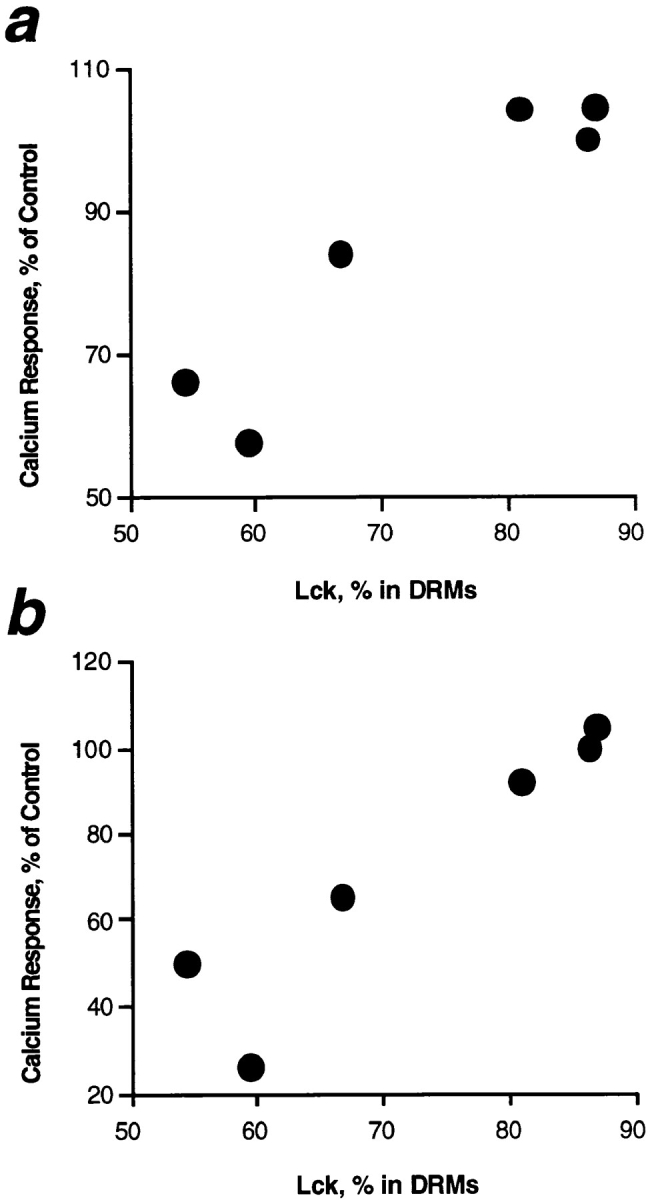
Polyunsaturated fatty acids (PUFAs) exert immunosuppressive effects, but the molecular alterations leading to T cell inhibition are not yet elucidated. Signal transduction seems to involve detergent-resistant membrane domains (DRMs) acting as functional rafts within the plasma membrane bilayer with Src family protein tyrosine kinases being attached to their cytoplasmic leaflet. Since DRMs include predominantly saturated fatty acyl moieties, we investigated whether PUFAs could affect T cell signaling by remodeling of DRMs. Jurkat T cells cultured in PUFA-supplemented medium showed a markedly diminished calcium response when stimulated via the transmembrane CD3 complex or glycosyl phosphatidylinositol (GPI)- anchored CD59. Immunofluorescence studies indicated that CD59 but not Src family protein tyrosine kinase Lck remained in a punctate pattern after PUFA enrichment. Analysis of DRMs revealed a marked displacement of Src family kinases (Lck, Fyn) from DRMs derived from PUFA-enriched T cells compared with controls, and the presence of Lck in DRMs strictly correlated with calcium signaling. In contrast, GPI-anchored proteins (CD59, CD48) and ganglioside GM1, both residing in the outer membrane leaflet, remained in the DRM fraction. In conclusion, PUFA enrichment selectively modifies the cytoplasmic layer of DRMs and this alteration could underlie the inhibition of T cell signal transduction by PUFAs.
Figures

References
-
- Ahmed SN, Brown DA, London E. On the origin of sphingolipid/ cholesterol rich detergent insoluble cell membranes: physiological concentrations of cholesterol and sphingolipid induce formation of a detergent insoluble, liquid ordered lipid phase in model membranes. Biochemistry. 1997;36:10944–10953. - PubMed
-
- Arni S, Ilangumaran S, van Echten G, Deckert, Sandhoff K, Poincelet M, Briol A, Rungger E, Brandle, Hoessli DC. Differential regulation of Src-family protein tyrosine kinases in GPI domains of T lymphocyte plasma membranes. Biochem Biophys Res Commun. 1996;225:801–807. - PubMed
-
- Belluzzi A, Brignola C, Campieri M, Pera A, Boschi S, Miglioli M. Effect of an enteric-coated fish-oil preparation on relapses in Crohn's disease. N Engl J Med. 1996;334:1557–1560. - PubMed
-
- Bligh EG, Dyer WJ. A rapid method for total lipid extraction and purification. Can J Biochem Physiol. 1959;37:911–917. - PubMed
-
- Bohuslav J, Cinek T, Hoř ejší V. Large, detergent-resistant complexes containing murine antigens Thy-1 and Ly-6 and protein tyrosine kinase p56lck. Eur J Immunol. 1993;23:825–831. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
