

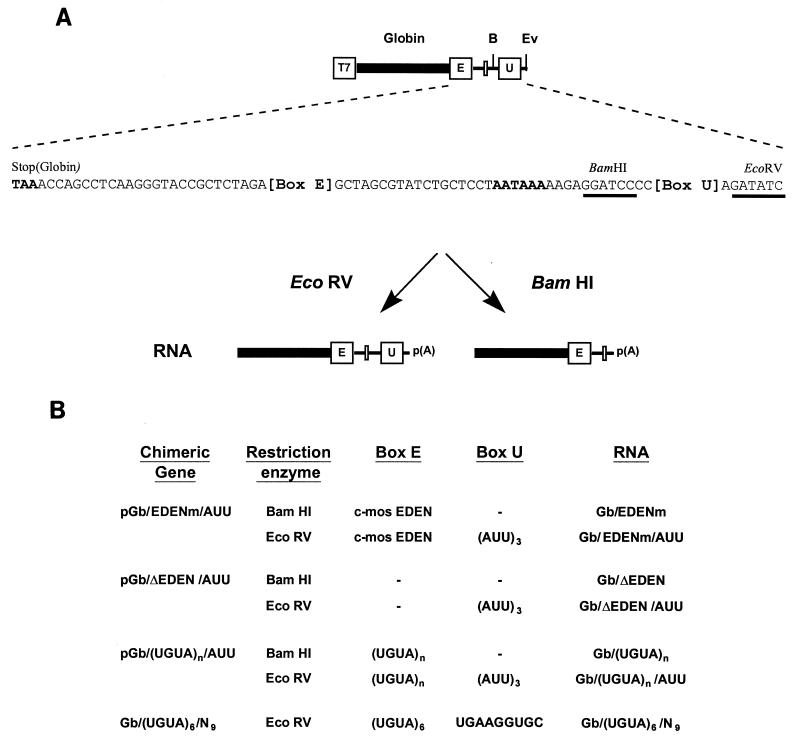
Embryo deadenylation element-dependent deadenylation is enhanced by a cis element containing AUU repeats
- PMID: 9819376
- PMCID: PMC109271
- DOI: 10.1128/MCB.18.12.6879
Embryo deadenylation element-dependent deadenylation is enhanced by a cis element containing AUU repeats
Abstract
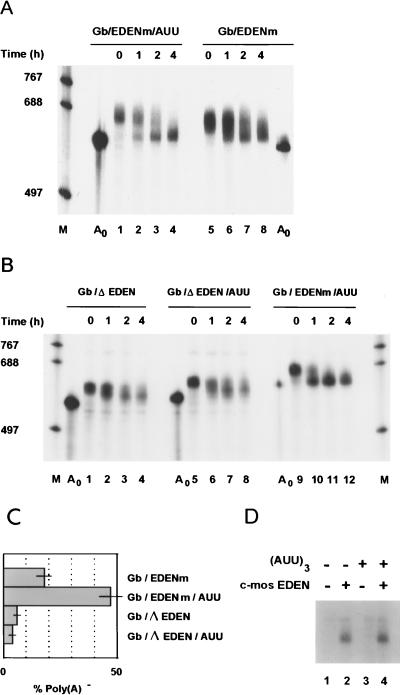
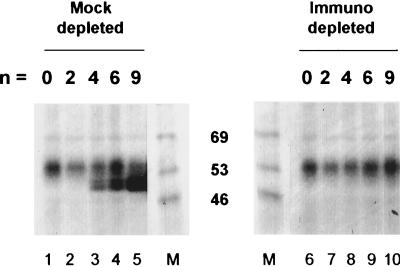
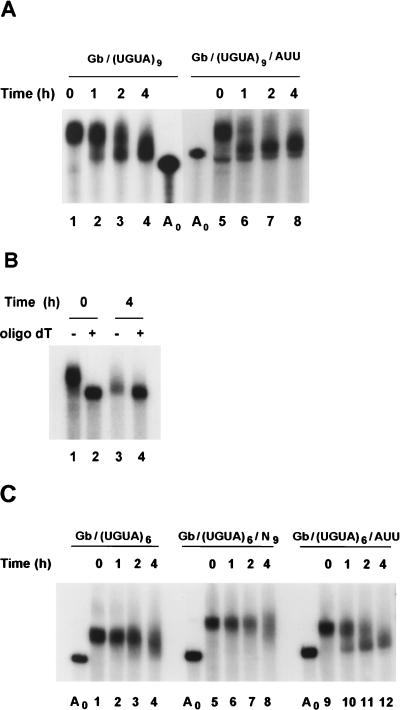
The deadenylation of maternal mRNAs in the Xenopus embryo is a sequence-specific process. One cis element that targets maternal mRNAs for deadenylation after fertilization is the embryo deadenylation element (EDEN). This element, composed of U/R repeats, is specifically bound by a protein, EDEN-BP. In the present study we show that the rate at which an RNA containing an EDEN is deadenylated can be increased by the presence of an additional cis element composed of three AUU repeats. This effect was observed for a natural EDEN (c-mos) and two synthetic EDENs. Hence, the enhancement of EDEN-dependent deadenylation conferred by the (AUU)3 motif is not due to an interaction with a particular EDEN sequence. Mutation of the (AUU)3 motif abrogated the enhancement of EDEN-dependent deadenylation. These data indicate that the rate at which a specific maternal mRNA is deadenylated in Xenopus embryos is probably defined by a cross talk between multiple cis elements.
Figures
References
-
- Audic Y, Omilli F, Osborne H B, Landais L. Design and use of easily made RNA size markers. BioTechniques. 1997;23:612–616. - PubMed
-
- Bouvet P, Omilli F, Arlot-Bonnemains Y, Legagneux V, Roghi C, Bassez T, Osborne H B. The deadenylation conferred by the 3′ untranslated region of a developmentally controlled mRNA in Xenopus embryos is switched to polyadenylation by deletion of a short sequence element. Mol Cell Biol. 1994;14:1893–1900. - PMC - PubMed
-
- Chevalier S, Blow J J. Cell cycle control of replication initiation in eukaryotes. Curr Opin Cell Biol. 1996;8:815–821. - PubMed
-
- Fox C A, Sheets M D, Wickens M. Poly(A) addition during maturation of frog oocytes: distinct nuclear and cytoplasmic activities and regulation by the sequence UUUUUAU. Genes Dev. 1989;3:2151–2162. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources