

The putative nucleic acid helicase Sen1p is required for formation and stability of termini and for maximal rates of synthesis and levels of accumulation of small nucleolar RNAs in Saccharomyces cerevisiae
- PMID: 9819377
- PMCID: PMC109272
- DOI: 10.1128/MCB.18.12.6885
The putative nucleic acid helicase Sen1p is required for formation and stability of termini and for maximal rates of synthesis and levels of accumulation of small nucleolar RNAs in Saccharomyces cerevisiae
Erratum in
- Mol Cell Biol 1999 Jul;19(7):5235
Abstract
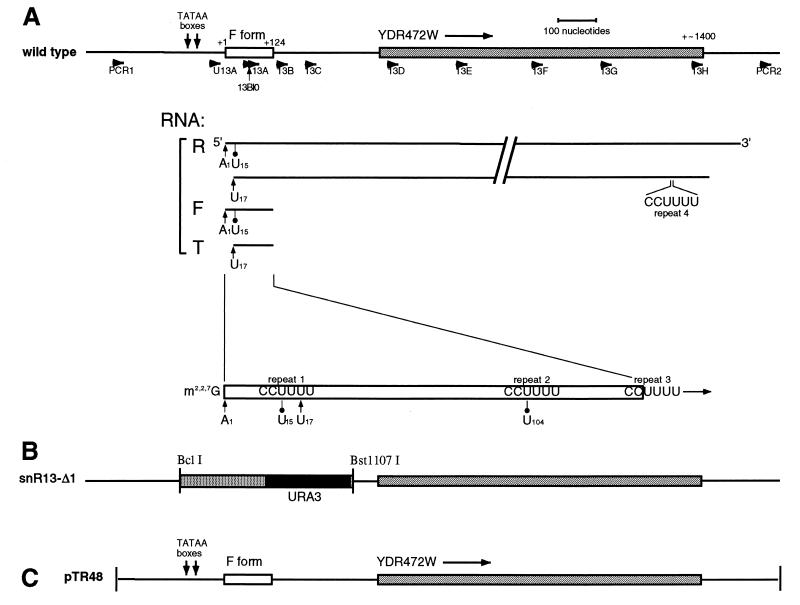
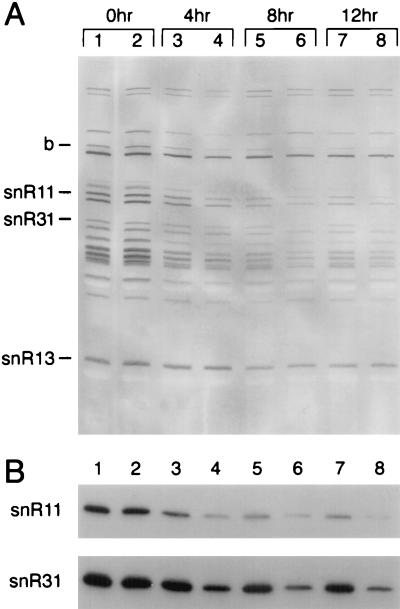
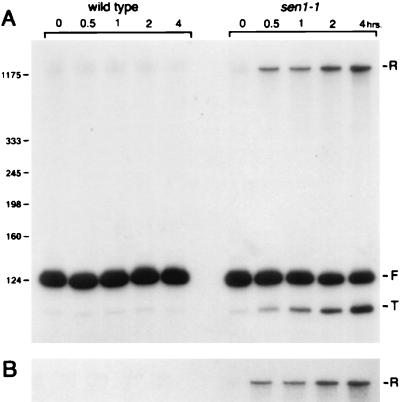
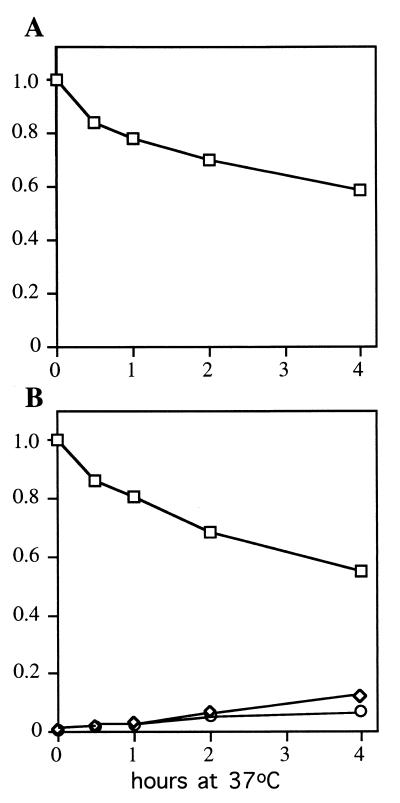
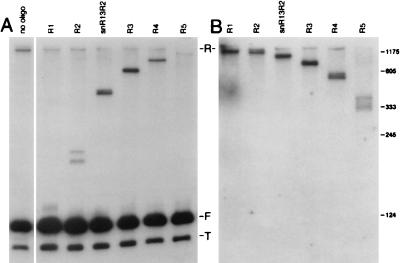
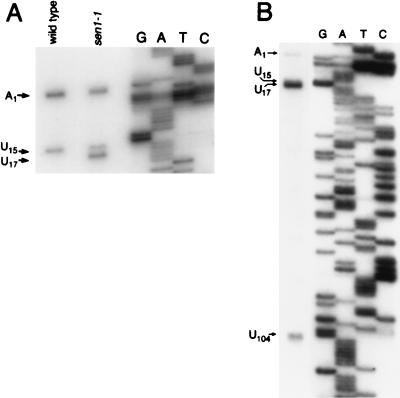
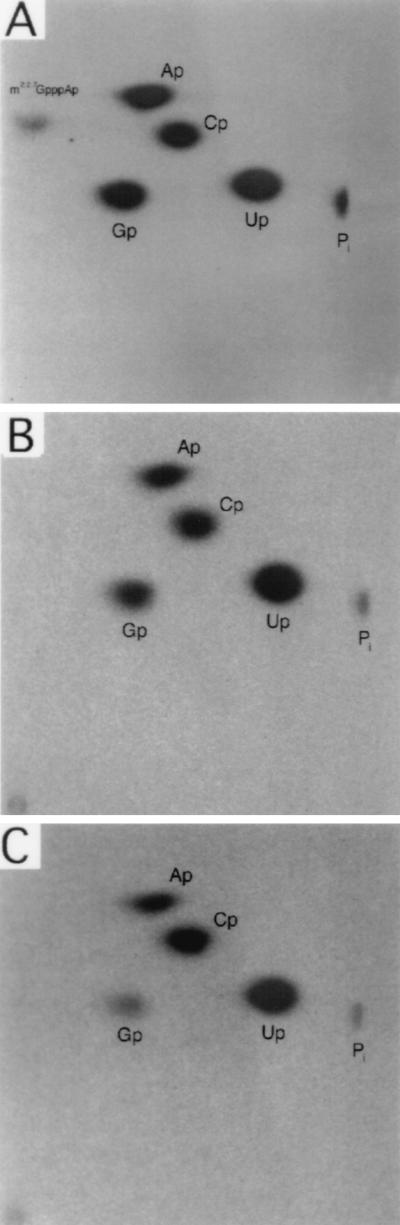
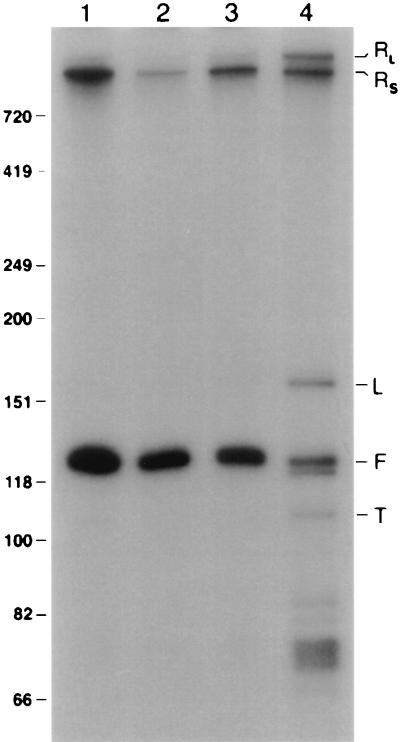
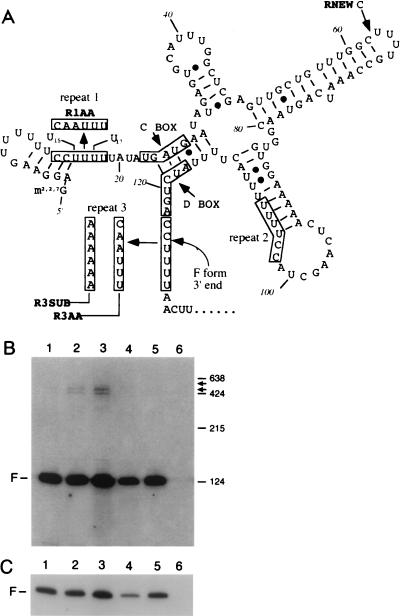
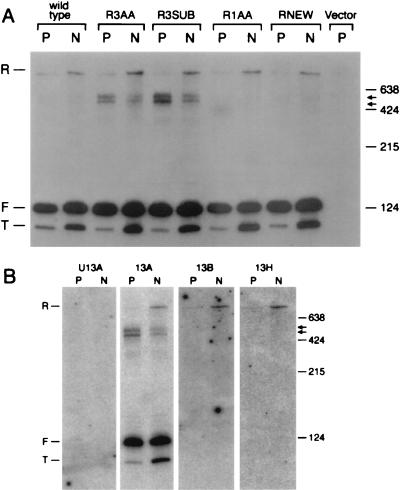
Sen1p from Saccharomyces cerevisiae is a nucleic acid helicase related to DEAD box RNA helicases and type I DNA helicases. The temperature-sensitive sen1-1 mutation located in the helicase motif alters the accumulation of pre-tRNAs, pre-rRNAs, and some small nuclear RNAs. In this report, we show that cells carrying sen1-1 exhibit altered accumulation of several small nucleolar RNAs (snoRNAs) immediately upon temperature shift. Using Northern blotting, RNase H cleavage, primer extension, and base compositional analysis, we detected three forms of the snoRNA snR13 in wild-type cells: an abundant TMG-capped 124-nucleotide (nt) mature form (snR13F) and two less abundant RNAs, including a heterogeneous population of approximately 1,400-nt 3'-extended forms (snR13R) and a 108-nt 5'-truncated form (snR13T) that is missing 16 nt at the 5' end. A subpopulation of snR13R contains the same 5' truncation. Newly synthesized snR13R RNA accumulates with time at the expense of snR13F following temperature shift of sen1-1 cells, suggesting a possible precursor-product relationship. snR13R and snR13T both increase in abundance at the restrictive temperature, indicating that Sen1p stabilizes the 5' end and promotes maturation of the 3' end. snR13F contains canonical C and D boxes common to many snoRNAs. The 5' end of snR13T and the 3' end of snR13F reside within C2U4 sequences that immediately flank the C and D boxes. A mutation in the 5' C2U4 repeat causes underaccumulation of snR13F, whereas mutations in the 3' C2U4 repeat cause the accumulation of two novel RNAs that migrate in the 500-nt range. At the restrictive temperature, double mutants carrying sen1-1 and mutations in the 3' C2U4 repeat show reduced accumulation of the novel RNAs and increased accumulation of snR13R RNA, indicating that Sen1p and the 3' C2U4 sequence act in a common pathway to facilitate 3' end formation. Based on these findings, we propose that Sen1p and the C2U4 repeats that flank the C and D boxes promote maturation of the 3' terminus and stability of the 5' terminus and are required for maximal rates of synthesis and levels of accumulation of mature snR13F.
Figures
References
-
- Balakin A G, Smith L, Fournier M J. The RNA world of the nucleolus: two major families of small RNAs defined by different box elements with related functions. Cell. 1996;86:823–834. - PubMed
-
- Brownlee G G. Determination of Sequences in RNA. New York, N.Y: American Elsevier Publishing; 1972.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials