

The T-cell oncogenic protein HOX11 activates Aldh1 expression in NIH 3T3 cells but represses its expression in mouse spleen development
- PMID: 9819390
- PMCID: PMC109285
- DOI: 10.1128/MCB.18.12.7030
The T-cell oncogenic protein HOX11 activates Aldh1 expression in NIH 3T3 cells but represses its expression in mouse spleen development
Abstract
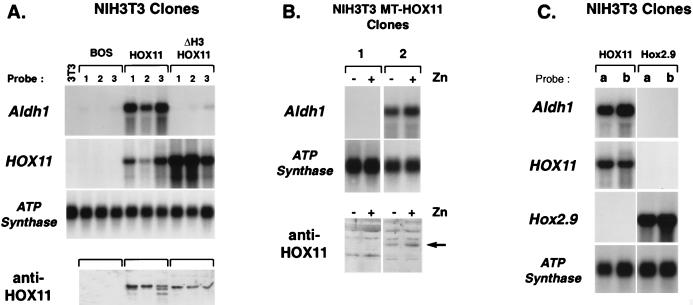
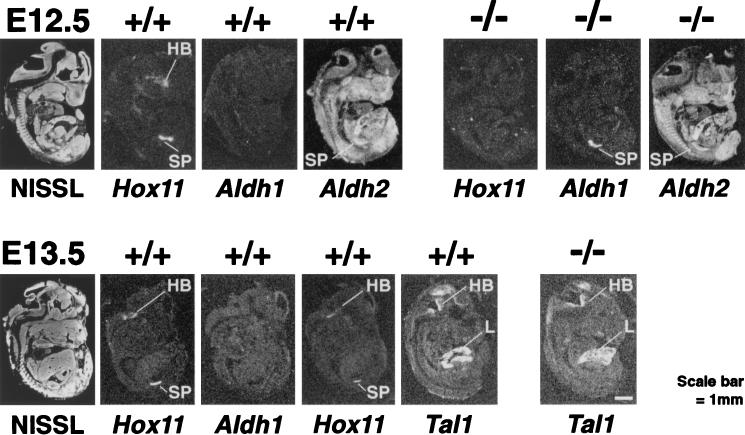
Hox11 is a homeobox gene essential for spleen formation in mice, since atrophy of the anlage of a developing spleen occurs in early embryonic development in Hox11 null mice. HOX11 is also expressed in a subset of T-cell acute leukemias after specific chromosomal translocations. Since the protein has a homeodomain and can activate transcription, it probably exerts at least some of its effects in vivo by regulation of target genes. Representational difference analysis has been used to isolate cDNA clones corresponding to mRNA species activated following stable expression of HOX11 in NIH 3T3 cells. The gene encoding the retinoic acid-synthesizing enzyme aldehyde dehydrogenase 1 (Aldh1), initially called Hdg-1, was found to be ectopically activated by HOX11 in this system. Study of Aldh1 gene expression during spleen development showed that the presence of Aldh1 mRNA inversely correlated with Hox11. Hox11 null mouse embryos have elevated Aldh1 mRNA in spleen primordia prior to atrophy, while Aldh1 seems to be repressed by Hox11 during organogenesis of the spleens of wild-type mice. This result suggests that expression of Aldh1 protein is negatively regulated by Hox11 and that abnormal expression of Aldh1 in Hox11 null mice may cause loss of splenic precursor cells by aberrant retinoic acid metabolism.
Figures


Similar articles
-
Specific homeodomain-DNA interactions are required for HOX11-mediated transformation.Blood. 2003 Jun 15;101(12):4966-74. doi: 10.1182/blood-2002-09-2857. Epub 2003 Feb 13. Blood. 2003. PMID: 12586625
-
The Hox11 gene is essential for cell survival during spleen development.Development. 1995 Sep;121(9):2909-15. doi: 10.1242/dev.121.9.2909. Development. 1995. PMID: 7555717
-
Hox11 is required to maintain normal Wt1 mRNA levels in the developing spleen.Dev Dyn. 2000 May;218(1):201-6. doi: 10.1002/(SICI)1097-0177(200005)218:1<201::AID-DVDY18>3.0.CO;2-R. Dev Dyn. 2000. PMID: 10822272
-
The promise of Hox11+ stem cells of the spleen for treating autoimmune diseases.Horm Metab Res. 2008 Feb;40(2):137-46. doi: 10.1055/s-2007-1022560. Horm Metab Res. 2008. PMID: 18283632 Review.
-
TALE homeoproteins as HOX11-interacting partners in T-cell leukemia.Leuk Lymphoma. 2000 Oct;39(3-4):241-56. doi: 10.3109/10428190009065824. Leuk Lymphoma. 2000. PMID: 11342305 Review.
Cited by
-
ALDH isozymes downregulation affects cell growth, cell motility and gene expression in lung cancer cells.Mol Cancer. 2008 Nov 24;7:87. doi: 10.1186/1476-4598-7-87. Mol Cancer. 2008. PMID: 19025616 Free PMC article.
-
TLX1/HOX11-induced hematopoietic differentiation blockade.Oncogene. 2007 Jun 14;26(28):4115-23. doi: 10.1038/sj.onc.1210185. Epub 2007 Jan 8. Oncogene. 2007. PMID: 17213805 Free PMC article.
-
Phosphorylation of HOX11/TLX1 on Threonine-247 during mitosis modulates expression of cyclin B1.Mol Cancer. 2010 Sep 16;9:246. doi: 10.1186/1476-4598-9-246. Mol Cancer. 2010. PMID: 20846384 Free PMC article.
-
Genes, proteins and complexes: the multifaceted nature of FHL family proteins in diverse tissues.J Cell Mol Med. 2010 Dec;14(12):2702-20. doi: 10.1111/j.1582-4934.2010.01176.x. J Cell Mol Med. 2010. PMID: 20874719 Free PMC article. Review.
-
The DN2 Myeloid-T (DN2mt) Progenitor is a Target Cell for Leukemic Transformation by the TLX1 Oncogene.J Bone Marrow Res. 2013 Feb 20;1:105. doi: 10.4172/2329-8820.1000105. J Bone Marrow Res. 2013. PMID: 25309961 Free PMC article.
References
-
- Ang H L, Duester G. Initiation of retinoid signalling in primitive streak mouse embryos: spatiotemporal expression patterns of receptors and metabolic enzymes for ligand synthesis. Dev Dyn. 1997;208:536–543. - PubMed
-
- Baer R. TAL1, TAL2 and LYL1: a family of basic helix-loop-helix proteins implicated in T cell acute leukaemia. Semin Cancer Biol. 1993;4:341–347. - PubMed
-
- Dear T N, Colledge W H, Carlton M B L, Lavenir I, Larson T, Smith A J H, Warren A J, Evans M J, Sofroniew M V, Rabbitts T H. The Hox11 gene is essential for cell survival during spleen development. Development. 1995;121:2909–2915. - PubMed
-
- Denhardt D T. A membrane-filter technique for the detection of complementary DNA. Biochem Biophys Res Commun. 1966;23:641–646. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous