

Effects of mitochondrial uncouplers on intracellular calcium, pH and membrane potential in rat carotid body type I cells
- PMID: 9824720
- PMCID: PMC2231310
- DOI: 10.1111/j.1469-7793.1998.819ba.x
Effects of mitochondrial uncouplers on intracellular calcium, pH and membrane potential in rat carotid body type I cells
Abstract
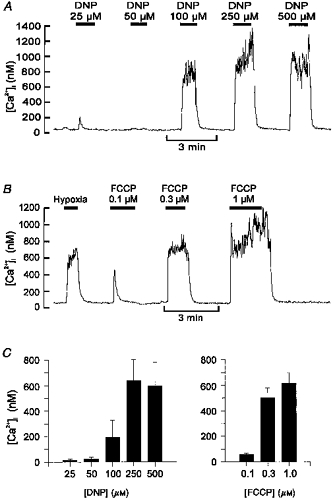
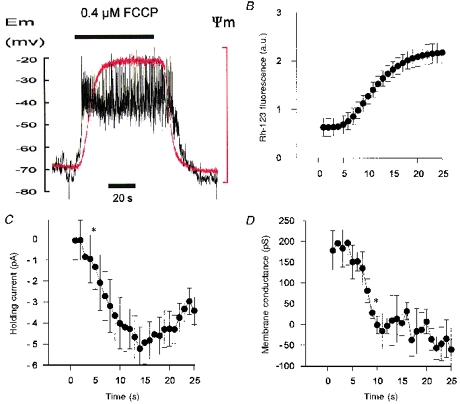
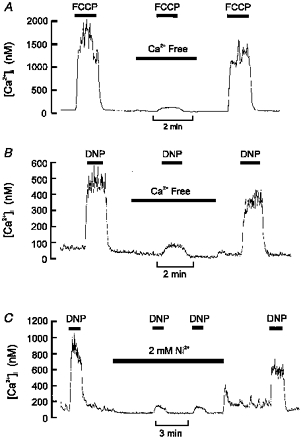
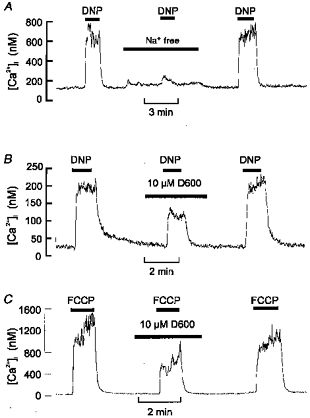
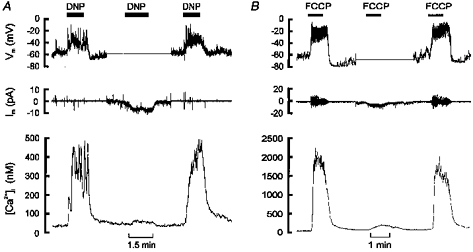
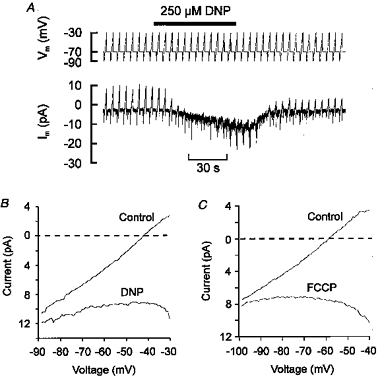
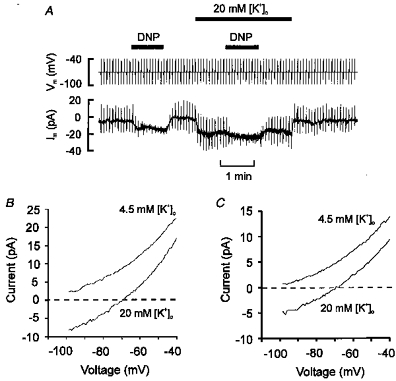
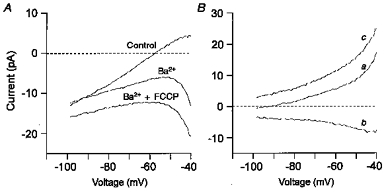
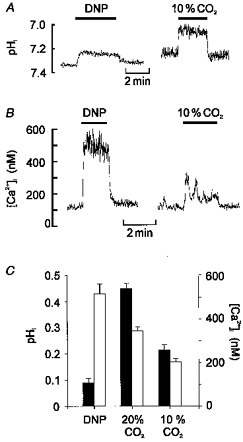
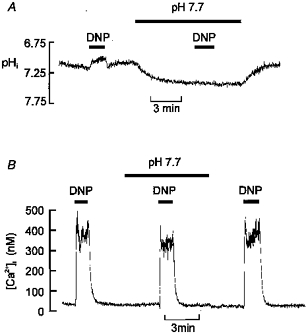
1. Mitochondrial uncouplers are potent stimulants of the carotid body. We have therefore investigated their effects upon isolated type I cells. Both 2,4-dinitrophenol (DNP) and carbonyl cyanide p-trifluoromethoxyphenyl hydrazone (FCCP) caused an increase in [Ca2+]i which was largely inhibited by removal of extracellular Ca2+ or Na+, or by the addition of 2 mM Ni2+. Methoxyverapamil (D600) also partially inhibited the [Ca2+]i response. 2. In perforated-patch recordings, the rise in [Ca2+]i coincided with membrane depolarization and was greatly reduced by voltage clamping the cell to -70 mV. Uncouplers also inhibited a background K+ current and induced a small inward current. 3. Uncouplers reduced pHi by 0.1 unit. Alkaline media diminished this acidification but had no effect on the [Ca2+]i response. 4. FCCP and DNP also depolarized type I cell mitochondria. The onset of mitochondrial depolarization preceded changes in cell membrane conductance by 3-4 s. 5. We conclude that uncouplers excite the carotid body by inhibiting a background K+ conductance and inducing a small inward current, both of which lead to membrane depolarization and voltage-gated Ca2+ entry. These effects are unlikely to be caused by cell acidification. The inhibition of background K+ current may be related to the uncoupling of oxidative phosphorylation.
Figures
References
-
- Anichcov SV, Belen'kii ML. Pharmacology of the Carotid Body Chemoreceptors. New York: Macmillan Publishing; 1963.
-
- Biscoe TJ, Duchen MR. The cellular basis of transduction in carotid chemoreceptors. American Journal of Physiology. 1990b;258:L271–278. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous