

Host factor I, Hfq, binds to Escherichia coli ompA mRNA in a growth rate-dependent fashion and regulates its stability
- PMID: 9826663
- PMCID: PMC24336
- DOI: 10.1073/pnas.95.24.14118
Host factor I, Hfq, binds to Escherichia coli ompA mRNA in a growth rate-dependent fashion and regulates its stability
Abstract
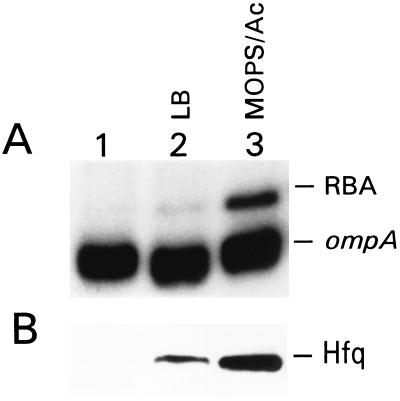
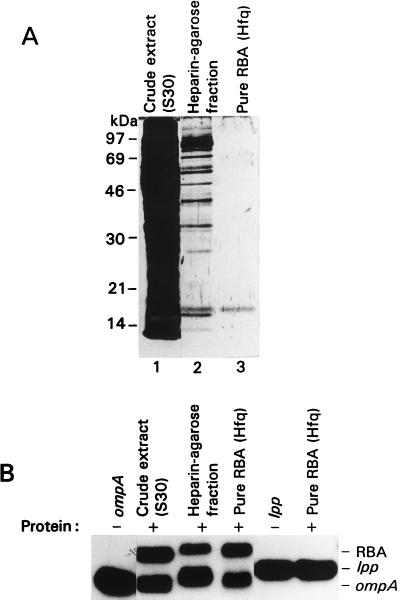
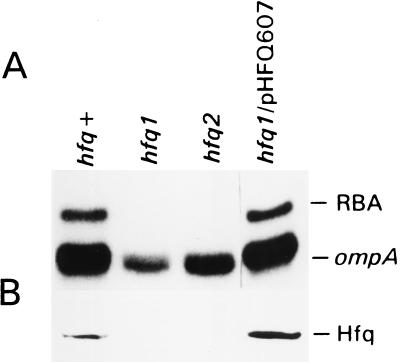
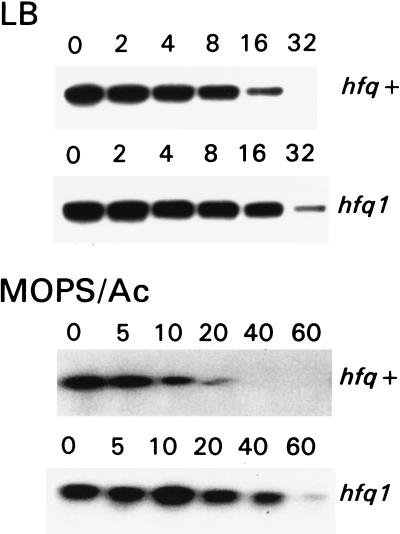
The stability of the ompA mRNA depends on the bacterial growth rate. The 5' untranslated region is the stability determinant of this transcript and the target of the endoribonuclease, RNase E, the key player of mRNA degradation. An RNA-binding protein with affinity for the 5' untranslated region ompA was purified and identified as Hfq, a host factor initially recognized for its function in phage Qbeta replication. The ompA RNA-binding activity parallels the amount of Hfq, which is elevated in bacteria cultured at slow growth rate, a condition leading to facilitated degradation of the ompA mRNA. In hfq mutant cells with a deficient Hfq gene product, the RNA-binding activity is missing, and analysis of the ompA mRNA showed that the growth-rate dependence of degradation is lost. Furthermore, the half-life of the ompA mRNA is prolonged in the mutant cells, irrespective of growth rate. Hfq has no affinity for the lpp transcript whose degradation, like that of bulk mRNA, is not affected by bacterial growth rate. Compatible with our results, we found that the intracellular concentration of RNase E and its associated degradosome components is independent of bacterial growth rate. Thus our results suggest a regulatory role for Hfq that specifically facilitates the ompA mRNA degradation in a growth rate-dependent manner.
Figures

References
-
- Lundberg U, Kaberdin V, von Gabain A. In: Manual of Industrial Microbiology and Biotechnology. Demain A L, Davies R M, Cohen G, Hershberg C L, Sherman D H, Willson R C, Wu J-H D, editors. Washington, DC: Am. Soc. Microbiol.; 1999. , in press.
-
- Nilsson G, Belasco J G, Cohen S N, von Gabain A. Nature (London) 1984;312:75–77. - PubMed
-
- Maaløe O. In: Biological Regulation and Development. Goldberger R F, editor. New York: Plenum; 1979. pp. 487–542.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
