

A functional homolog of a yeast tRNA splicing enzyme is conserved in higher eukaryotes and in Escherichia coli
- PMID: 9826666
- PMCID: PMC24339
- DOI: 10.1073/pnas.95.24.14136
A functional homolog of a yeast tRNA splicing enzyme is conserved in higher eukaryotes and in Escherichia coli
Abstract
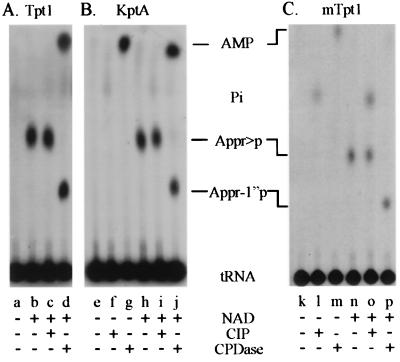
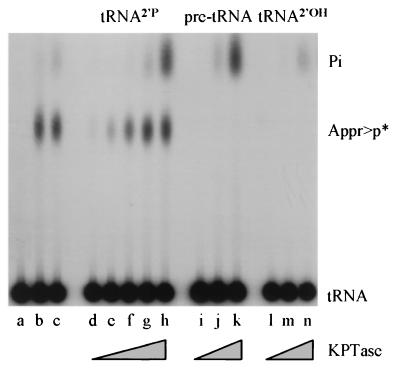
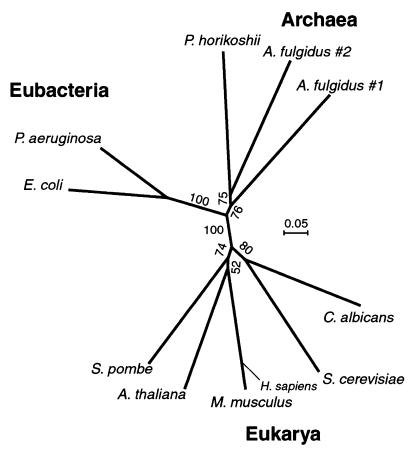
tRNA splicing in the yeast Saccharomyces cerevisiae requires an endonuclease to excise the intron, tRNA ligase to join the tRNA half-molecules, and 2'-phosphotransferase to transfer the splice junction 2'-phosphate from ligated tRNA to NAD, producing ADP ribose 1"-2" cyclic phosphate (Appr>p). We show here that functional 2'-phosphotransferases are found throughout eukaryotes, occurring in two widely divergent yeasts (Candida albicans and Schizosaccharomyces pombe), a plant (Arabidopsis thaliana), and mammals (Mus musculus); this finding is consistent with a role for the enzyme, acting in concert with ligase, to splice tRNA or other RNA molecules. Surprisingly, functional 2'-phosphotransferase is found also in the bacterium Escherichia coli, which does not have any known introns of this class, and does not appear to have a ligase that generates junctions with a 2'-phosphate. Analysis of the database shows that likely members of the 2'-phosphotransferase family are found also in one other bacterium (Pseudomonas aeruginosa) and two archaeal species (Archaeoglobus fulgidus and Pyrococcus horikoshii). Phylogenetic analysis reveals no evidence for recent horizontal transfer of the 2'-phosphotransferase into Eubacteria, suggesting that the 2'-phosphotransferase has been present there since close to the time that the three kingdoms diverged. Although 2'-phosphotransferase is not present in all Eubacteria, and a gene disruption experiment demonstrates that the protein is not essential in E. coli, the continued presence of 2'-phosphotransferase in Eubacteria over large evolutionary times argues for an important role for the protein.
Figures

References
-
- Xu M Q, Kathe S D, Goodrich-Blair H, Nierzwicki-Bauer S A, Shub D A. Science. 1990;250:1566–1570. - PubMed
-
- Reinhold-Hurek B, Shub D A. Nature (London) 1992;357:173–176. - PubMed
-
- Ferat J L, Michel F. Nature (London) 1993;364:358–361. - PubMed
-
- Belfort M, Weiner A. Cell. 1997;89:1003–1006. - PubMed
-
- Abelson J, Trotta C R, Li H. J Biol Chem. 1998;273:12685–12688. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
