

Explosive invasion of plant mitochondria by a group I intron
- PMID: 9826685
- PMCID: PMC24358
- DOI: 10.1073/pnas.95.24.14244
Explosive invasion of plant mitochondria by a group I intron
Abstract
Group I introns are mobile, self-splicing genetic elements found principally in organellar genomes and nuclear rRNA genes. The only group I intron known from mitochondrial genomes of vascular plants is located in the cox1 gene of Peperomia, where it is thought to have been recently acquired by lateral transfer from a fungal donor. Southern-blot surveys of 335 diverse genera of land plants now show that this intron is in fact widespread among angiosperm cox1 genes, but with an exceptionally patchy phylogenetic distribution. Four lines of evidence-the intron's highly disjunct distribution, many incongruencies between intron and organismal phylogenies, and two sources of evidence from exonic coconversion tracts-lead us to conclude that the 48 angiosperm genera found to contain this cox1 intron acquired it by 32 separate horizontal transfer events. Extrapolating to the over 13,500 genera of angiosperms, we estimate that this intron has invaded cox1 genes by cross-species horizontal transfer over 1,000 times during angiosperm evolution. This massive wave of lateral transfers is of entirely recent occurrence, perhaps triggered by some key shift in the intron's invasiveness within angiosperms.
Figures
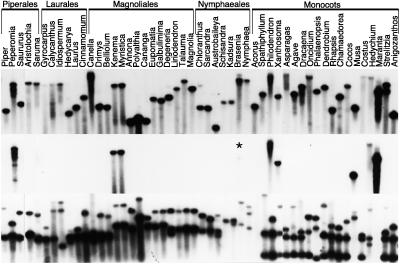
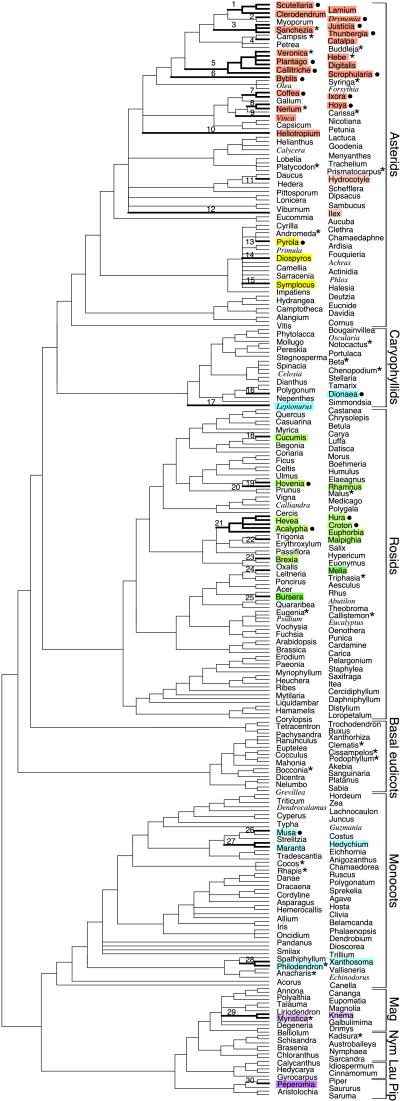
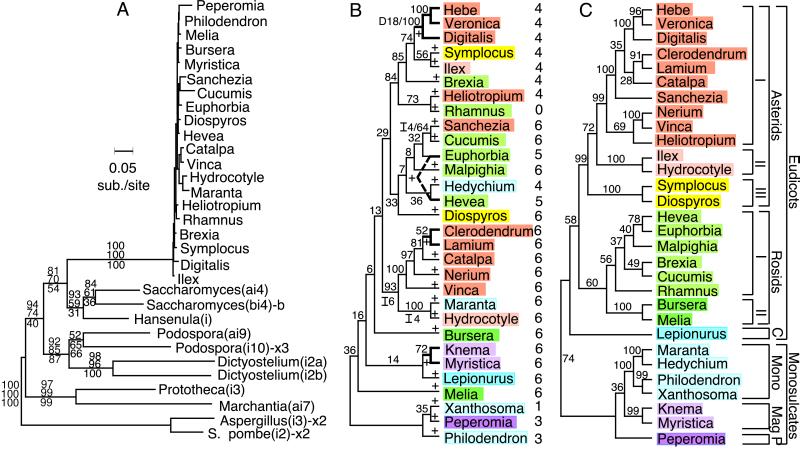

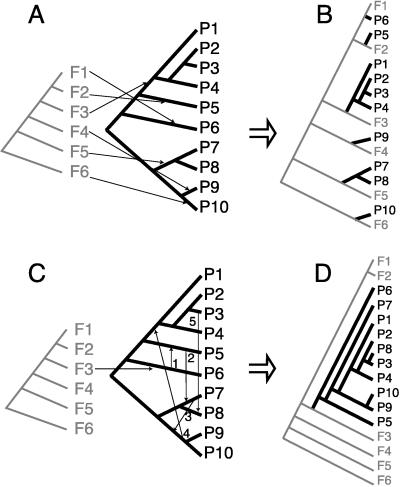
Comment in
-
Mass migration of a group I intron: promiscuity on a grand scale.Proc Natl Acad Sci U S A. 1998 Nov 24;95(24):14003-5. doi: 10.1073/pnas.95.24.14003. Proc Natl Acad Sci U S A. 1998. PMID: 9826641 Free PMC article. Review. No abstract available.
References
-
- Dujon B. Gene. 1989;82:91–114. - PubMed
-
- Lambowitz A M, Belfort M. Annu Rev Biochem. 1993;62:587–622. - PubMed
-
- Belfort M, Perlman P S. J Biol Chem. 1995;270:30237–30240. - PubMed
-
- Delahodde A, Goguel V, Becam A M, Creusot F, Perea J, Banroques J, Jacq C. Cell. 1989;56:431–441. - PubMed
-
- Wenzlau J M, Saldanha R J, Butow R A, Perlman P S. Cell. 1989;56:421–430. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
