

Rpb4, a subunit of RNA polymerase II, enables the enzyme to transcribe at temperature extremes in vitro
- PMID: 9829926
- PMCID: PMC107702
- DOI: 10.1128/JB.180.23.6187-6192.1998
Rpb4, a subunit of RNA polymerase II, enables the enzyme to transcribe at temperature extremes in vitro
Abstract
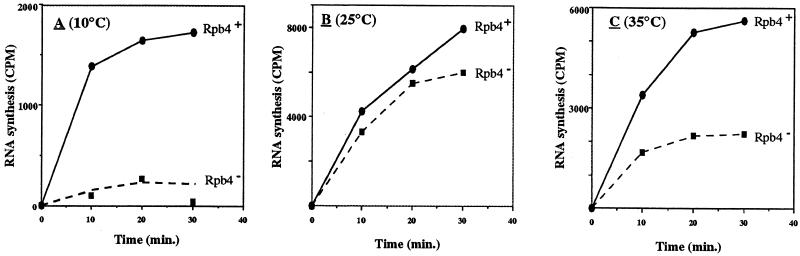
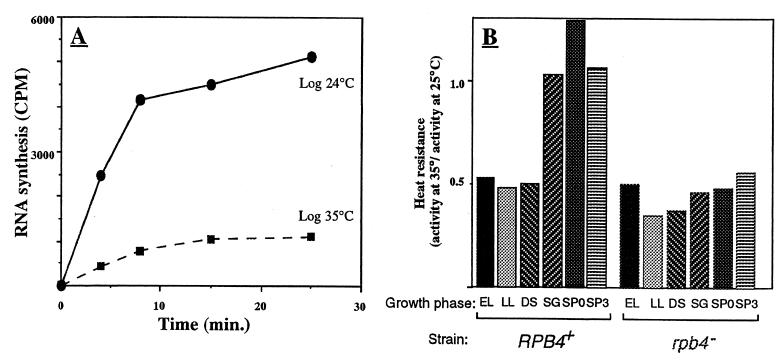
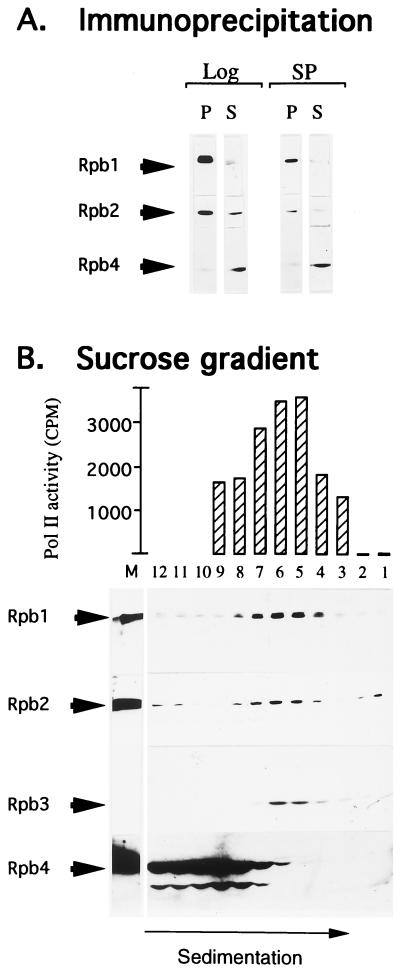
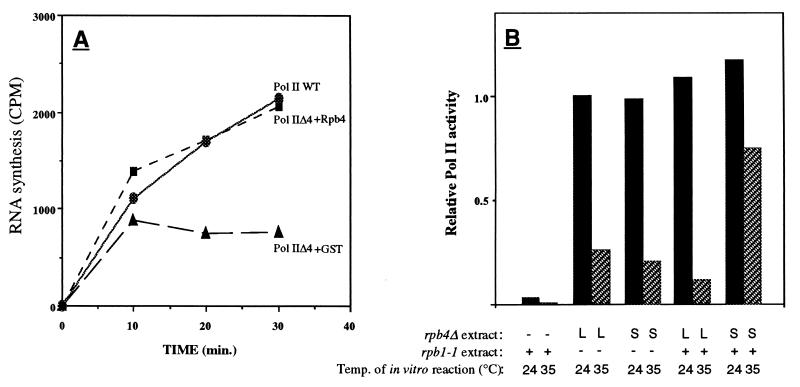
Rpb4 is a subunit of Saccharomyces cerevisiae RNA polymerase II (Pol II). It associates with the polymerase preferentially in stationary phase and is essential for some stress responses. Using the promoter-independent initiation and chain elongation assay, we monitored Pol II enzymatic activity in cell extracts. We show here that Rpb4 is required for the polymerase activity at temperature extremes (10 and 35 degreesC). In contrast, at moderate temperature (23 degreesC) Pol II activity is independent of Rpb4. These results are consistent with the role previously attributed to Rpb4 as a subunit whose association with Pol II helps Pol II to transcribe during extreme temperatures. The enzymatic inactivation of Pol II lacking Rpb4 at the nonoptimal temperature was prevented by the addition of recombinant Rpb4 produced in Escherichia coli prior to the in vitro reaction assay. This finding suggests that modification of Rpb4 is not required for its functional association with the other Pol II subunits. Sucrose gradient and immunoprecipitation experiments demonstrated that Rpb4 is present in the cell in excess over the Pol II complex during all growth phases. Nevertheless, the rescue of Pol II activity at the nonoptimal temperature by Rpb4 is possible only when cell extracts are obtained from postlogarithmic cells, not from logarithmically growing cells. This result suggests that Pol II molecules should be modified in order to recruit Rpb4; the portion of the modified Pol II molecules is small during logarithmic phase and becomes predominant in stationary phase.
Figures
References
-
- Acker J, de Graaff M, Cheynel I, Khazak V, Kedinger C, Vigneron M. Interactions between the human RNA polymerase II subunits. J Biol Chem. 1997;272:16815–16821. - PubMed
-
- Austrias F J, Meredith G D, Poglitsch C L, Kornberg R D. Two conformation of RNA polymerase II revealed by electron crystallography. J Mol Biol. 1997;272:536–540. - PubMed
-
- Choder M. A general topoisomerase I-dependent transcriptional repression in the stationary phase in yeast. Genes Dev. 1991;5:2315–2326. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
