

Regional crypt function in rat large intestine in relation to fluid absorption and growth of the pericryptal sheath
- PMID: 9831728
- PMCID: PMC2269061
- DOI: 10.1111/j.1469-7793.1999.211af.x
Regional crypt function in rat large intestine in relation to fluid absorption and growth of the pericryptal sheath
Abstract
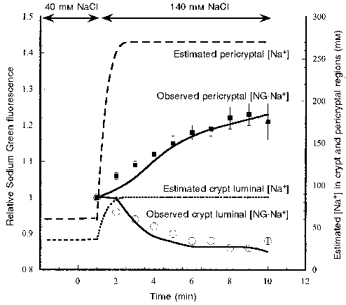
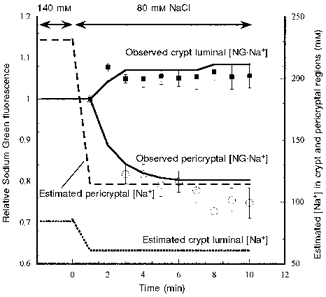
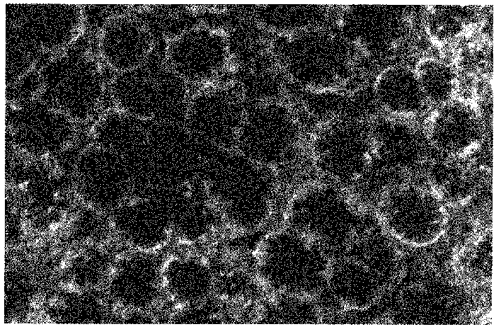
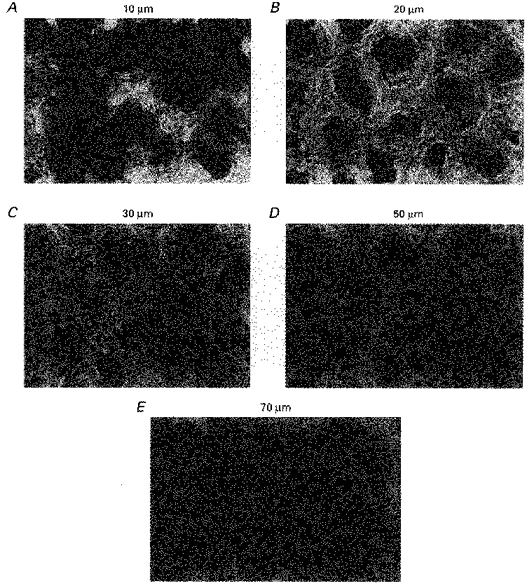
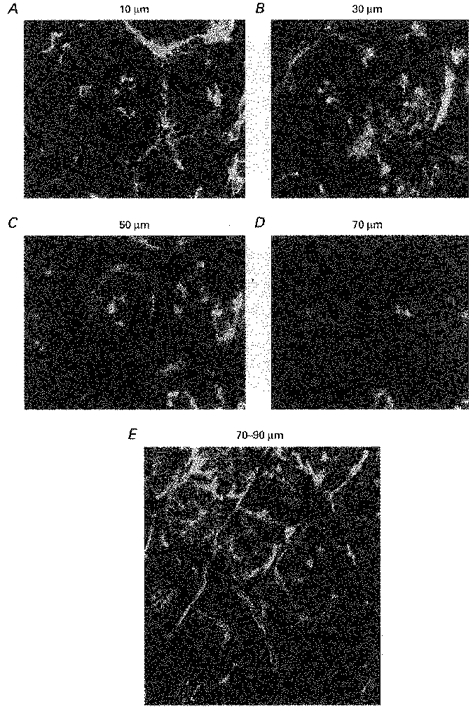
1. Confocal microscopic studies of rat colonic mucosa showed that the pericryptal sheath surrounding distal colonic crypts is an effective barrier both to dextran and NaCl movement, whereas no such structure surrounds the caecal crypts. 2. The distal colonic pericryptal barrier was functionally demonstrated by accumulation of Sodium Green within the pericryptal space. After exposure to benzamil, Sodium Green accumulation was decreased. Fluorescein isocyanate-labelled dextran (FITC dextran; molecular mass 10000 Da) was accumulated in the crypt lumens and pericryptal spaces. Both dextran and Sodium Green accumulation were absent from the pericryptal zone surrounding caecal crypts. 3. Low dietary Na+ intake raised rat plasma aldosterone and stimulated distal pericryptal sheath growth and adhesiveness as shown by increased amounts of F-actin, smooth muscle actin, beta-catenin and E-cadherins in the pericryptal zone. It also raised the capacity of the distal colon to dehydrate against a high luminal hydraulic resistance. This linkage indicates that trophic effects on the colon resulting from a low Na+ diet are not confined solely to effects on transepithelial Na+ transport, but are observed in the pericryptal sheath. 4. A computer model of crypt function confirms that a pericryptal sheath with low permeability to NaCl is an essential component of the crypt dehydrating mechanism.
Figures

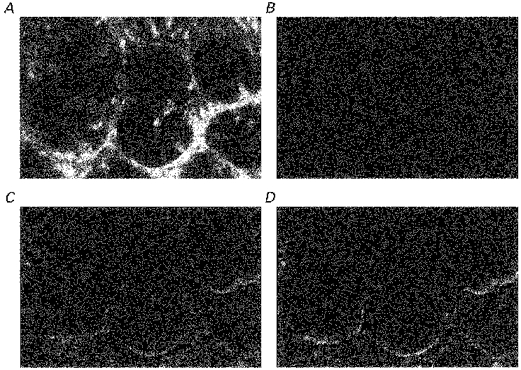

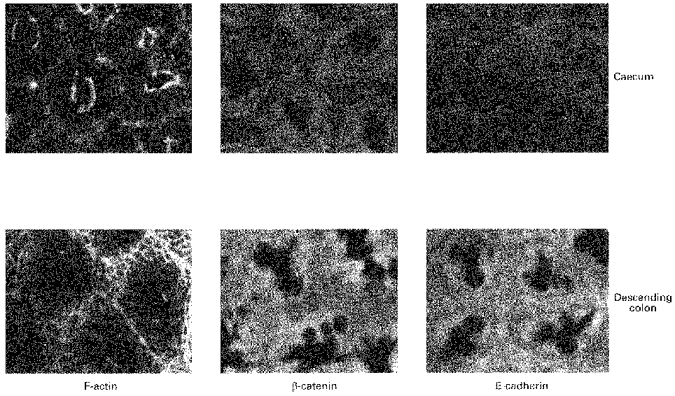
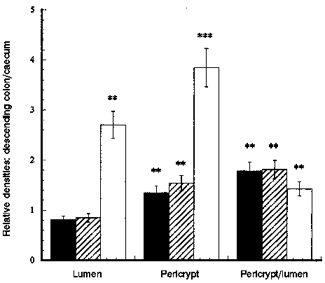

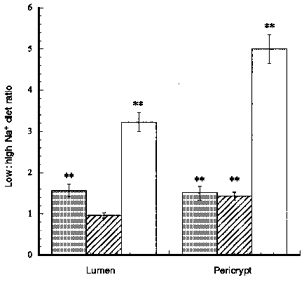
 ) and β-catenin (□) distribution in descending colonic crypts of rat fed on high and low Na+ diets Each column shows mean values from tissues from 3 rats each fed comtemporaneously on high or low Na+ diets. Four regions of interest in the crypt and pericryptal regions from each tissue were evaluated. **P < 0.01 low Na+vs. high Na+ diet; Student's two-tailed t test.
) and β-catenin (□) distribution in descending colonic crypts of rat fed on high and low Na+ diets Each column shows mean values from tissues from 3 rats each fed comtemporaneously on high or low Na+ diets. Four regions of interest in the crypt and pericryptal regions from each tissue were evaluated. **P < 0.01 low Na+vs. high Na+ diet; Student's two-tailed t test.



References
-
- Abayasekara DRE, Onyezili NI, Whitehouse BJ, Laird SM, Vinson GP. Effects of chronic corticotrophin treatment on aldosterone metabolism in the rat. Journal of Endocrinology. 1993;137:445–455. - PubMed
-
- Araki K, Furuya Y, Kobayashi M, Matsuura K, Ogata T, Isozaki H. Comparison of mucosal microvasculature between the proximal and distal human colon. Journal of Electron Microscopy. 1996;45:202–206. - PubMed
-
- Bleakman D, Naftalin RJ. Hypertonic fluid absorption from rat descending colon in vitro. American Journal of Physiology. 1990;258:G377–390. - PubMed
-
- Bridges RJ, Cragoe EJ, Jr, Frizzell RA, Benos DJ. Inhibition of colonic Na+ transport by amiloride analogues. American Journal of Physiology. 1989;256:C67–74. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources