

Potassium currents in freshly dissociated uterine myocytes from nonpregnant and late-pregnant rats
- PMID: 9834143
- PMCID: PMC2229446
- DOI: 10.1085/jgp.112.6.737
Potassium currents in freshly dissociated uterine myocytes from nonpregnant and late-pregnant rats
Abstract
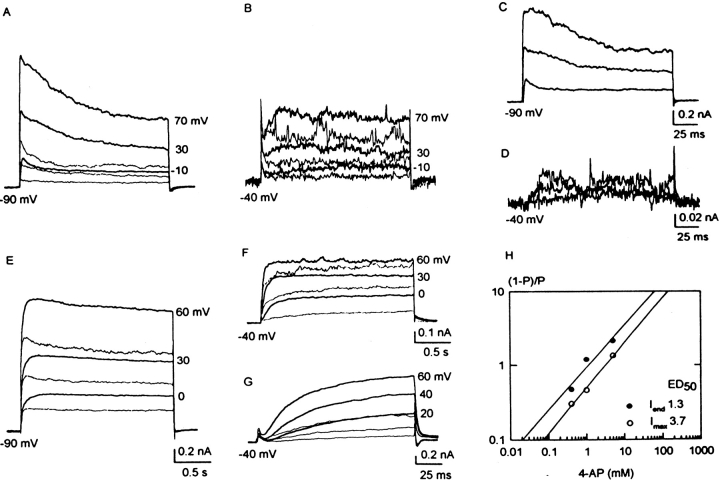
In freshly dissociated uterine myocytes, the outward current is carried by K+ through channels highly selective for K+. Typically, nonpregnant myocytes have rather noisy K+ currents; half of them also have a fast-inactivating transient outward current (ITO). In contrast, the current records are not noisy in late pregnant myocytes, and ITO densities are low. The whole-cell IK of nonpregnant myocytes respond strongly to changes in [Ca2+]o or changes in [Ca2+]i caused by photolysis of caged Ca2+ compounds, nitr 5 or DM-nitrophene, but that of late-pregnant myocytes respond weakly or not at all. The Ca2+ insensitivity of the latter is present before any exposure to dissociating enzymes. By holding at -80, -40, or 0 mV and digital subtractions, the whole-cell IK of each type of myocyte can be separated into one noninactivating and two inactivating components with half-inactivation at approximately -61 and -22 mV. The noninactivating components, which consist mainly of iberiotoxin-susceptible large-conductance Ca2+-activated K+ currents, are half-activated at 39 mV in nonpregnant myocytes, but at 63 mV in late-pregnant myocytes. In detached membrane patches from the latter, identified 139 pS, Ca2+-sensitive K+ channels also have a half-open probability at 68 mV, and are less sensitive to Ca2+ than similar channels in taenia coli myocytes. Ca2+-activated K+ currents, susceptible to tetraethylammonium, charybdotoxin, and iberiotoxin contribute 30-35% of the total IK in nonpregnant myocytes, but <20% in late-pregnant myocytes. Dendrotoxin-susceptible, small-conductance delayed rectifier currents are not seen in nonpregnant myocytes, but contribute approximately 20% of total IK in late-pregnant myocytes. Thus, in late-pregnancy, myometrial excitability is increased by changes in K+ currents that include a suppression of the ITO, a redistribution of IK expression from large-conductance Ca2+-activated channels to smaller-conductance delayed rectifier channels, a lowered Ca2+ sensitivity, and a positive shift of the activation of some large-conductance Ca2+-activated channels.
Figures













Similar articles
-
Comparison of voltage-dependent potassium currents in rat pyramidal neurons acutely isolated from hippocampal regions CA1 and CA3.J Neurophysiol. 1995 Nov;74(5):1982-95. doi: 10.1152/jn.1995.74.5.1982. J Neurophysiol. 1995. PMID: 8592191
-
Ca(2+)-activated K+ channels in pregnant rat myometrium: modulation by a beta-adrenergic agent.Am J Physiol. 1992 Nov;263(5 Pt 1):C1049-56. doi: 10.1152/ajpcell.1992.263.5.C1049. Am J Physiol. 1992. PMID: 1359792
-
A novel large-conductance Ca(2+)-activated potassium channel and current in nerve terminals of the rat neurohypophysis.J Physiol. 1992 Nov;457:47-74. doi: 10.1113/jphysiol.1992.sp019364. J Physiol. 1992. PMID: 1284313 Free PMC article.
-
Characterization of two distinct depolarization-activated K+ currents in isolated adult rat ventricular myocytes.J Gen Physiol. 1991 May;97(5):973-1011. doi: 10.1085/jgp.97.5.973. J Gen Physiol. 1991. PMID: 1865177 Free PMC article.
-
Calcium influx via L- and N-type calcium channels activates a transient large-conductance Ca2+-activated K+ current in mouse neocortical pyramidal neurons.J Neurosci. 2003 May 1;23(9):3639-48. doi: 10.1523/JNEUROSCI.23-09-03639.2003. J Neurosci. 2003. PMID: 12736335 Free PMC article.
Cited by
-
Role of potassium channels in female reproductive system.Obstet Gynecol Sci. 2020 Sep;63(5):565-576. doi: 10.5468/ogs.20064. Epub 2020 Aug 25. Obstet Gynecol Sci. 2020. PMID: 32838485 Free PMC article.
-
Voltage-gated K+ currents in freshly isolated myocytes of the pregnant human myometrium.J Physiol. 1999 Aug 1;518 ( Pt 3)(Pt 3):769-81. doi: 10.1111/j.1469-7793.1999.0769p.x. J Physiol. 1999. PMID: 10420013 Free PMC article.
-
Pregnancy switches adrenergic signal transduction in rat and human uterine myocytes as probed by BKCa channel activity.J Physiol. 2000 Apr 15;524 Pt 2(Pt 2):339-52. doi: 10.1111/j.1469-7793.2000.t01-1-00339.x. J Physiol. 2000. PMID: 10766916 Free PMC article.
-
Potassium channels and uterine function.Semin Cell Dev Biol. 2007 Jun;18(3):332-9. doi: 10.1016/j.semcdb.2007.05.008. Epub 2007 May 24. Semin Cell Dev Biol. 2007. PMID: 17596977 Free PMC article. Review.
-
A role for voltage-gated, but not Ca2+-activated, K+ channels in regulating spontaneous contractile activity in myometrium from virgin and pregnant rats.Br J Pharmacol. 2006 Apr;147(7):815-24. doi: 10.1038/sj.bjp.0706644. Br J Pharmacol. 2006. PMID: 16415906 Free PMC article.
References
-
- Amman, D. 1986. Ion Selective Microelectrodes. Springer Verlag GmbH & Co. Berlin, Germany. 109–207.
-
- Blatz AL, Magleby KL. Single apamine-blocked Ca2+-activated K+channels of small conductance in cultured rat skeletal muscle. Nature. 1986;323:718–720. - PubMed
-
- Chandy, K.G., and G.A. Gutman. 1995. Voltage-gated potassium channel genes. In Handbook of Receptors and Channels, Ligand and Voltage-Gated Ion Channels. R.A. North, editor. CRC Press. Boca Raton, FL. 1–71.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
