

Molecular cloning and functional expression in lactobacillus plantarum 80 of xylT, encoding the D-xylose-H+ symporter of Lactobacillus brevis
- PMID: 9835554
- PMCID: PMC90914
- DOI: 10.1128/AEM.64.12.4720-4728.1998
Molecular cloning and functional expression in lactobacillus plantarum 80 of xylT, encoding the D-xylose-H+ symporter of Lactobacillus brevis
Abstract
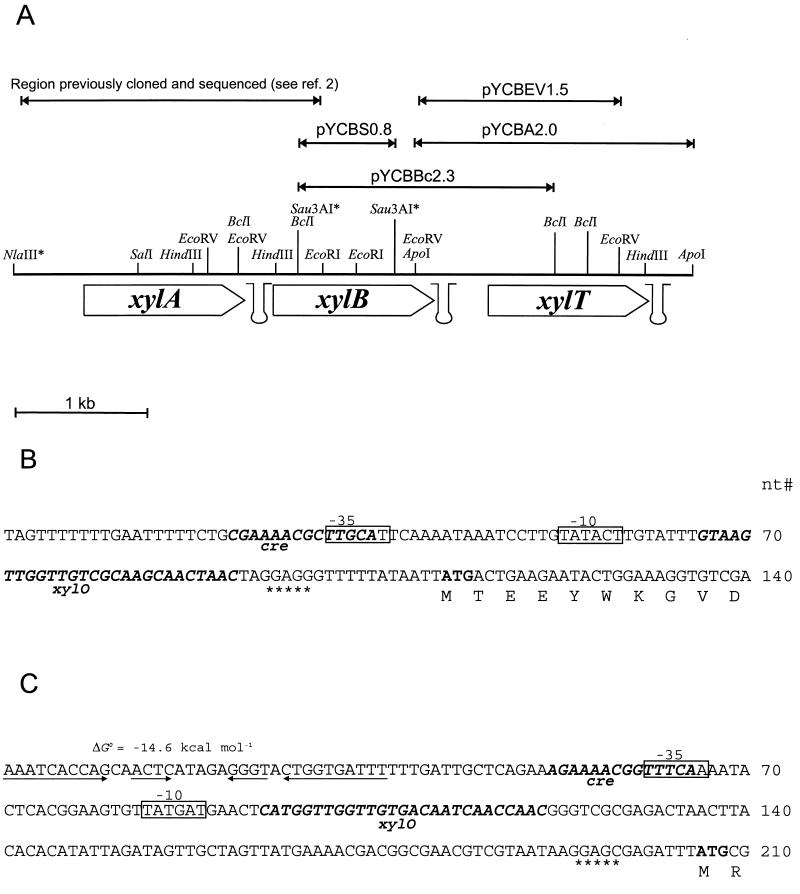
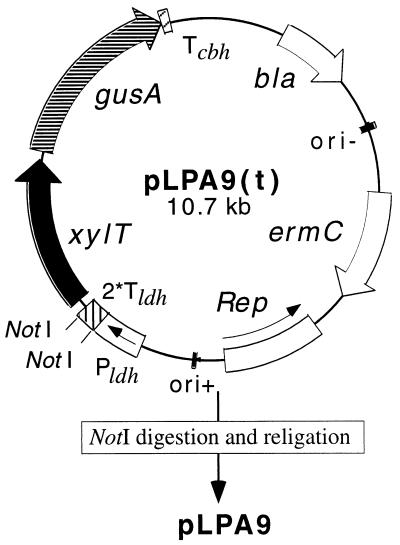
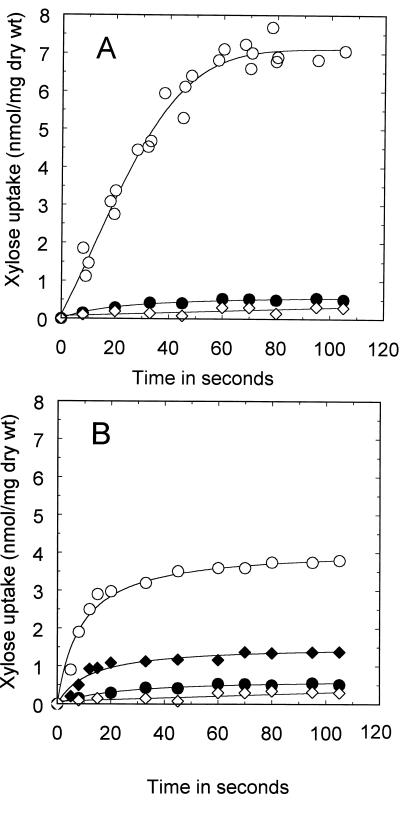
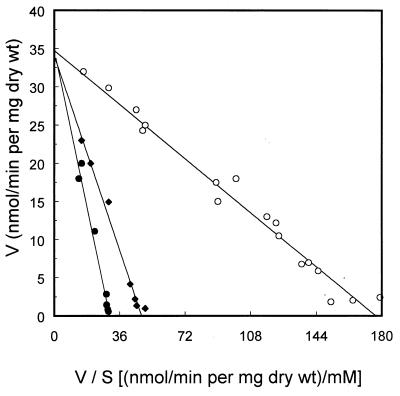
A 3-kb region, located downstream of the Lactobacillus brevis xylA gene (encoding D-xylose isomerase), was cloned in Escherichia coli TG1. The sequence revealed two open reading frames which could code for the D-xylulose kinase gene (xylB) and another gene (xylT) encoding a protein of 457 amino acids with significant similarity to the D-xylose-H+ symporters of E. coli, XylE (57%), and Bacillus megaterium, XylT (58%), to the D-xylose-Na+ symporter of Tetragenococcus halophila, XylE (57%), and to the L-arabinose-H+ symporter of E. coli, AraE (60%). The L. brevis xylABT genes showed an arrangement similar to that of the B. megaterium xylABT operon and the T. halophila xylABE operon. Southern hybridization performed with the Lactobacillus pentosus xylR gene (encoding the D-xylose repressor protein) as a probe revealed the existence of a xylR homologue in L. brevis which is not located with the xyABT locus. The existence of a functional XylR was further suggested by the presence of xylO sequences upstream of xylA and xylT and by the requirement of D-xylose for the induction of D-xylose isomerase, D-xylulose kinase, and D-xylose transport activities in L. brevis. When L. brevis was cultivated in a mixture of D-glucose and D-xylose, the D-xylose isomerase and D-xylulose kinase activities were reduced fourfold and the D-xylose transport activity was reduced by sixfold, suggesting catabolite repression by D-glucose of D-xylose assimilation. The xylT gene was functionally expressed in Lactobacillus plantarum 80, a strain which lacks proton motive force-linked D-xylose transport activity. The role of the XylT protein was confirmed by the accumulation of D-xylose in L. plantarum 80 cells, and this accumulation was dependent on the proton motive force generated by either malolactic fermentation or by the metabolism of D-glucose. The apparent affinity constant of XylT for D-xylose was approximately 215 microM, and the maximal initial velocity of transport was 35 nmol/min per mg (dry weight). Furthermore, of a number of sugars tested, only 6-deoxy-D-glucose inhibited the transport of D-xylose by XylT competitively, with a Ki of 220 microM.
Figures

References
-
- Altschul S F, Gish W, Miller W, Myers E W, Lipman D J. Basic local alignment search tool. J Mol Biol. 1990;215:403–410. - PubMed
-
- Bor Y-C, Moraes S-P, Lee S-P, Crosby W L, Sinskey A J, Batt C A. Cloning and sequencing the Lactobacillus brevis gene encoding xylose isomerase. Gene. 1992;114:127–131. - PubMed
-
- Callens M, Kerstens-Hildersion H, van Opstal O, de Bruine C K. Catalytic properties of d-xylose isomerase from Streptomyces violaceoruber. Enzyme Microb Technol. 1986;8:696–700.
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
